
Energieumwandelnde biologische Prozesse sind in der Regel membrangebunden. Dadurch unterscheiden sie sich grundsätzlich von einfachen enzymkatalysierten Reaktionen, die in Lösung ablaufen können. Energiegewinn konnte es also erst geben, nachdem die funktionelle Struktur membranumgebener Vesikel (Zellen oder zumindest Zellvorstufen) vorhanden war. Im Verlauf der Photosynthese und der Atmungskette entstehen Protonengradienten; die Membran wird durch sie polarisiert, und die im Gradienten gespeicherte Energie wird zur Bildung einer energiereichen chemischen Bindung, einer Phosphorylierung (ADP + Pi > ATP) verwendet. In Form der energiereichen Bindung kann die Zelle die Energie für andere Prozesse nutzen.
Modellversuche ergaben, daß eine Photophosphorylierung (d.h. eine Phosphorylierung unter Ausnutzung von Lichtenergie) unter abiotischen Bedingungen mit geringer Ausbeute auch in Abwesenheit von Membranen, aber in Anwesenheit von Molekülen ablaufen kann, die es schon sehr früh nach Bildung der Erdkruste gegeben haben konnte (s. Simulationsexperimente von S. L. MILLER in Thema Wie ist Leben entstanden ?):
(W. S. BRINIGAR, D. B. KNAFF, J. H. WANG, 1967).
Primitive Prokaryoten können ihren Energiebedarf durch Elektronenentzug aus anorganischem Material decken (Chemolitho-Autotrophie), weiter fortentwickelte durch Vergärung organischen Materials (Heterotrophie).
Ein einfacher Fall einer Umwandlung von Lichtenergie in chemische Energie wurde bei einem Archaebacterium (Halobacterium halobium) analysiert. An dessen "Photosynthese" ist nur ein einziges Protein beteiligt; es fehlt eine Elektronentransportkette, die für die fortentwickelten energiegewinnenden Mechanismen typisch ist. Die Energieausbeute ist bei Halobacterium außerordentlich niedrig.
Eine Photosynthese in dem Sinne wie sie uns heute bei allen grünen Pflanzen, bei Blaualgen und photosynthetisierenden Bakterien begegnet, beruht auf der Ausnutzung von Lichtenergie (ursprünglich mit hohem UV-Anteil), d.h., einer Umsetzung eines Photonenflusses in einen Elektronenfluß und der Ausbildung von Elektronentransportketten.
Photoautotrophe Organismen enthalten meist Chlorophyll a (oder ein ihm verwandtes Derivat wie das Bakterienchlorophyll a). Im Gegensatz zu den Häminen enthält der Porphyrinanteil der Chlorophylle Magnesiumionen anstelle von Ferro- / Ferriionen als Zentralatom. Vermutlich erfolgte der Wechsel zu Magnesium, weil der dadurch entstandene Komplex die vorhandene Lichtmenge und Lichtqualität, emittiert von der Sonne besser nutzen konnte.
Unterschiedlich sind die Membranproteine, an die Chlorophyll a gebunden ist sowie die Zusammensetzung der übrigen "akzessorischen" Pigmente. Man geht davon aus, daß die Uratmosphäre reduzierend war. Als Elektronendonatoren kamen daher Substanzen wie Wasserstoff, Schwefelwasserstoff und andere Schwefelverbindungen in Betracht.
 Die Nutzung
von Wasser als Protonen- und Elektronendonatoren findet man erstmals bei
den Blaualgen (Cyanophyceae, Cyanobacteria). Als Folge ihrer Photosyntheseaktivität
geriet freier Sauerstoff in die Atmosphäre, dessen Konzentration sich
von ursprünglich nahe Null auf den heutigen Wert von ca. 20 Prozent
einpendelte. Damit änderten sich die Selektionsbedingungen für
die Organismen grundlegend. Diejenigen gewannen die Überhand, die
sich vor dem Sauerstoff schützen und ihn schließlich auch nutzen
konnten.
Die Nutzung
von Wasser als Protonen- und Elektronendonatoren findet man erstmals bei
den Blaualgen (Cyanophyceae, Cyanobacteria). Als Folge ihrer Photosyntheseaktivität
geriet freier Sauerstoff in die Atmosphäre, dessen Konzentration sich
von ursprünglich nahe Null auf den heutigen Wert von ca. 20 Prozent
einpendelte. Damit änderten sich die Selektionsbedingungen für
die Organismen grundlegend. Diejenigen gewannen die Überhand, die
sich vor dem Sauerstoff schützen und ihn schließlich auch nutzen
konnten.
Die Zunahme der Sauerstoffmenge führte zur Ausbildung einer Ozonschicht in der Stratosphäre, und damit zur Verschiebung der spektralen Zusammensetzung des die Erdoberfläche erreichenden Lichts. Kurzwelliges Licht (UV) wurde weitgehend weggefiltert. Für die Organismen ergab sich damit die Notwendigkeit, in steigendem Maße Licht des sichtbaren Bereichs zu nutzen, und in Folge dieser Ereignisse änderte sich die Pigmentzusammensetzung der Photosysteme.
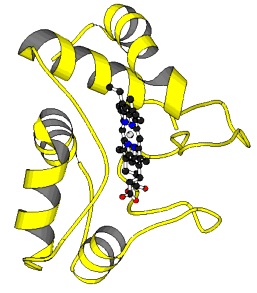
Zum weiteren Verständnis der Entwicklung energiegewinnender Prozesse muß die Evolution der Elektronentransportketten betrachtet werden. Zu den Gemeinsamkeiten von Photosynthese und Atmungskette gehört die Beteiligung gleichartiger Proteine: Ferredoxin, Cytochrom c, Cytochrom b u.a.
Die Cytochrome b6 (aus Plastiden) und b (aus Mitochondrien) sind aufgrund topologischer Verwandtschaft miteinander homologisierbar. Das Cytochrom b6 entspricht in der Abfolge von hydrophoben und hydrophilen Aminosäuresequenzabschnitten einer entsprechenden Sequenz in der N-terminalen Hälfte des Cytochroms b. (W. R. WIDGER, W. A. CRAMER, R. G. HERRMANN, A. TREBST: Purdue University, Universität München und Universität Bochum, 1984). Die Homologisierbarkeit der entsprechenden Cytochrome ist ein weiterer stichhaltiger Beweis dafür, daß Photosynthese und Atmungskette auf ein gleiches Reaktionsmuster zurückgeführt werden können.
Zu einem ähnlichen Schluß gelangt man durch Vergleiche der Topologie der am Elektronentransport beteiligten Komplexe. Die Atmungskette ist demnach eine Variante der Photosynthese; sie vervollkommnete sich parallel zur Zunahme an freiem Sauerstoff.

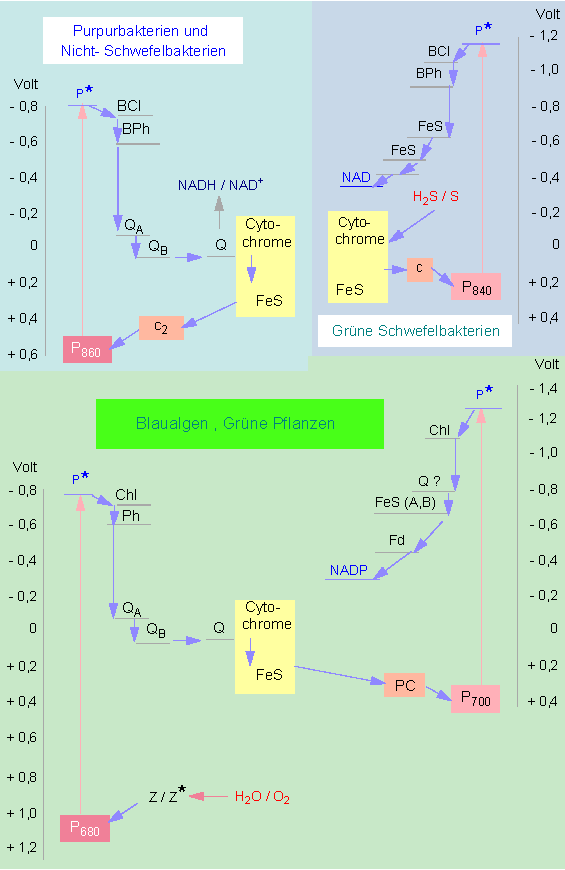
Die Photosysteme I und II entstanden ursprünglich unabhängig voneinander (es sei zunächst noch dahingestellt, ob auch sie untereinander homologisierbar sind); durch Zusammenschluß entstaNd ein System (Z-Schema der Photosynthese), das Lichtenergie wesentlich effizienter nutzen konnte als jedes der Teilsysteme für sich allein.
|
|