
Erst in den letzten Jahren begann man, sich auch für die pflanzlichen Mitochondrien zu interessieren, doch stehen uns aufgrund der Analyse von Mitochondrien aus anderen Quellen, z.B. Pilzen (Saccharomyces cerevisiae, Neurospora crassa), Ciliaten und Vertebraten genügend Daten zur Verfügung, um uns ein generelles Bild von ihnen machen zu können. Mit den Chloroplasten haben die Mitochondrien eine Reihe von Gemeinsamkeiten:
![]() sie enthalten genetische Information (mtDNS).
sie enthalten genetische Information (mtDNS).
![]() sie enthalten eine eigene Proteinsynthesemaschinerie (eigene Ribosomen,
tRNS, u.a),
sie enthalten eine eigene Proteinsynthesemaschinerie (eigene Ribosomen,
tRNS, u.a),
![]() sie besitzen neben einer äußeren eine innere Membran, an
der Energieumwandlungen stattfinden.
sie besitzen neben einer äußeren eine innere Membran, an
der Energieumwandlungen stattfinden.
Nach der Endosymbiontenhypothese sind beide Organelltypen aus Prokaryoten hervorgegangen, die eine Symbiose mit primitiven eukaryotischen Zellen eingegangen sind.
Alle im vorangegangenen Abschnitt aufgeworfenen Fragen stellen sich auch hier. Nur ein geringer Teil der in Mitochondrien benötigten Proteine wird dort selbst synthetisiert, die meisten stammen aus dem Cytosol. Das Mitochondriengenom des Menschen ist sequenziert worden, das mtDNS-Molekül enthält 16 5691 Basenpaare (S. ANDERSON et al., 1981). Auf höhere Pflanzen lassen sich demnach viele der an Vertebraten und Hefe-mtDNS gewonnenen Einzelerkenntnisse nicht ausdehnen. Die mtDNS höherer Pflanzen ist extrem komplex. Die Molekulargewichte schwanken zwischen 200 000 und 2 400 000. In einer Pflanze findet man zudem ein weites Spektrum verschiedener Moleküle. Neben großen linearen Molekülen kommen unterschiedlich große zirkuläre vor. Viele der Moleküle sind unvollständig, und Rekombinationsereignisse scheinen häufig zu sein. An anderer Stelle wurde darauf hingewiesen, daß der genetische Code in Mitochondrien anders als sonst gelesen wird.
Besondere Aufmerksamkeit wurde in den letzten Jahren dem Phänomen der cytoplasmatisch vererbten Pollensterilität (CMS: cytoplasmic male sterility) geschenkt, da eine solche - im übrigen bei höheren Pflanzen verbreitete - Hemmung der Pollenentwicklung zur Herstellung von Hybridsaatgut genutzt wird. Beim Mais kennt man mindestens drei CMS-Linien: C, T und S. Nur Pflanzen mit dem Cytoplasma des Typs N (=normal) produzieren fertilen Pollen. Es zeigte sich, daß die mtDNS des C-Plasmas sich gegenüber der von N durch zahlreiche Umstrukturierungen auszeichnet. Es entstehen auf diese Weise hybride Gene, z.B. ein Gen, das aus einem Anteil eines ATPase-Gens und einem des Cytochrom-Oxydase-Gens besteht. Die mtDNS der T-Linie ist noch wesentlich komplexer; gegenüber der von N zeichnet sie sich durch einen extrem hohen Umstrukturierungsgrad aus. In den Mitochondrien der S-Linie wurden zwei lineare Plasmide gefunden. Plasmide sind zusätzliche kleine DNS-Moleküle, die für Bakterien typisch sind und die sich in den letzten Jahren als unentbehrliche Hilfsmittel der Gentechnik bewährt haben. Ihr Nachweis in eukaryotischen Zellen gelang bei Pilzen (auch dort sind sie vornehmlich, aber nicht ausschließlich, in Mitochondrien lokalisiert) und in höheren Pflanzen (das hier zitierte Beispiel ist kein Einzelfall). Es sieht so aus, als würden die in den Mitochondrien der CMS-Linien gebildeten Hybridproteine in die innere Mitochondrienmembran eingelagert und deren Funktion beeinträchtigen.
Eine Reihe bestimmter Enzymaktivitäten findet sich simultan sowohl im Cytosol als auch in den Mitochondrien (und ggf. auch in den Chloroplasten). Die dafür benötigten Enzyme haben die gleiche Spezifität, unterscheiden sich aber in ihrer Struktur (Aminosäurezusammensetzung). Damit ist sichergestellt, daß einer der Typen in das eine Organell, der andere in das zweite transportiert werden, während der dritte im Cytosol verbleibt. Enzyme mit gleicher Aktivität und Spezifität, doch anderer Struktur und damit auch anderer Reaktionskinetik, bezeichnet man als Isoenzyme. Wenngleich man von den meisten solcher Isoenzyme die genauen proteinchemischen Daten nicht kennt, weiß man doch, daß sie proteinchemisch verschieden sind, denn sie verhalten sich unterschiedlich in der Gelelektrophorese und/oder der isoelektrischen Fokussierung.
Auch Mitochondrien entstehen durch Teilung auseinander. Während in tierischen Zellen ausschließlich die Wechselwirkung zwischen Kerngenom und Mitochondriengenom zu betrachten ist, sind wir in Zellen grüner Pflanzen mit einem Dreiecksverhältnis von Kerngenom-Chloroplastengenom-Mitochondrien-Genom konfrontiert. Vor allem die innere mitochondriale Membran ist selbst für niedermolekulare Metaboliten nur selektiv passierbar. In ihr enthaltene Carrier bedingen die Selektivität und steuern die Durchflußrate.
Die wichtigste Leistung der Mitochondrien ist und bleibt aber die Atmungskettenphosphorylierung oder Oxydative Phosphorylierung. Sie besteht aus zwei funktionellen Abschnitten, einmal der Elektronentransportkette und zum anderen der ATP-Bildung, also der eigentlichen Phosphorylierungsreaktion. Die ATP-Bildung wird durch eine ATP-Synthetase katalysiert, die ein Bestandteil der inneren Mitochondrienmembran ist. Wie schon dargelegt wurde und wie bei der Besprechung der Photophosphorylierung erneut betont werden wird, kann dieses Enzym nur dann arbeiten, wenn die Membran intakt, d.h. absolut protonenundurchlässig ist und zwei Kompartimente voneinander trennt, so daß sich ein Protonengradient über die Membran hinweg aufbauen kann. Dieser Gradient ist die treibende Kraft, die zur Phosphorylierungsreaktion benötigt wird.
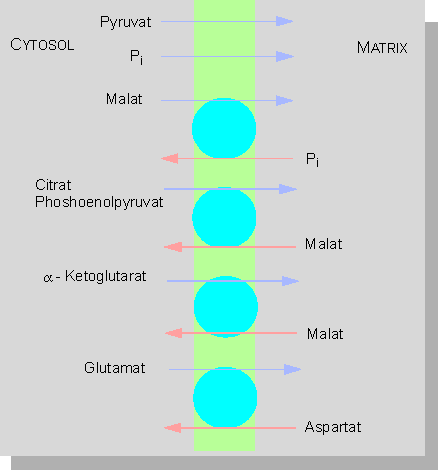
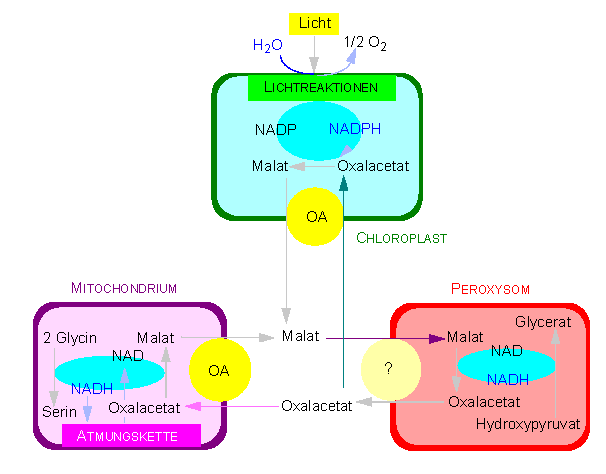
Malat-Oxalacetat-Shuttle: Ein charakteristisches Merkmal pflanzlicher Zellen. Er dient dem intrazellulären Transport von Redoxäquivalenten. Die entscheidende Bedeutung dabei hat der Oxalacetat-Translokator (OA) (Nach H. W. HELDT und U. T. FLÜGGE, 1986)
|
|