

Das Prinzip der hierarchischen Klassifikation von Pflanzen (und anderen Organismen) ist der nebenstehenden Abbildung zu entnehmen. Die Zugehörigkeit der einzelnen Einheiten richtet sich nach abgestuften Ähnlichkeiten. Dadurch entsteht ein System unterschiedlicher Hierarchieebenen. Je umfassender eine Gruppe ist, desto geringer sind die Gemeinsamkeiten zwischen ihren Mitgliedern. Andererseits ist das Vorhandensein klar definierter Abgrenzungen zwischen ihnen das wichtigste Kriterium zur Untergliederung der Gruppe. Überall dort, wo fließende Übergänge vorhanden sind, wäre eine Grenzziehung willkürlich.
Trägt man den Prozentsatz der Ähnlichkeiten gegen die Zahl der Einheiten auf, erhält man ein Dendrogramm, aus dem die Hierarchie der Rangstufen ablesbar ist.
Die Unterscheidung zwischen Arten und übergeordneten Gruppen kann durch eine Folge von Ja/Nein-Entscheidungen herbeigeführt werden. Ein Dendrogramm ist im einfachsten Fall dichotom aufgebaut. Nach einem Schlüssel dieser Art, erstmals von J. B. de LAMARCK vorgeschlagen, arbeiten die meisten Bestimmungsbücher. Trotz gelegentlicher Einwände und mancher zweifelhafter Fälle hat sich dieses Prinzip bewährt. In der vorangegangenen Abbildung sind bereits Benennungen der Rangstufen aufgenommen worden, die in der biologischen Systematik üblich sind.Taxonomische (systematische) Gruppen, gleich welcher Kategorie oder Rangstufe, nennt man Taxa (sing. Taxon).
Die wissenschaftlichen Namen aller taxonomischen Gruppen werden gewöhnlich der lateinischen oder griechischen Sprache entnommen. Entstammen sie anderen Sprachen, müssen sie wie lateinische Namen behandelt werden (Fuchsia, z.B., ist die Latinisierung des Namens FUCHS).
Artnamen bestehen aus zwei Elementen (binäre Nomenklatur), dem Gattungsnamen und dem nachfolgenden Epithet, durch das der Artname festgelegt wird. Dem Namen folgt der Name des Erstbeschreibers (Autors). Die Namen bekannter Systematiker der vergangenen Jahrhunderte werden oft abgekürzt, L. steht für LINNÉ. Benennungen aus der Zeit vor LINNÉ werden nicht berücksichtigt. Unterarten werden durch einen dritten, ebenfalls lateinischen Namen gekennzeichnet, der den Artnamen ergänzt. Zum Beispiel Poa trivialis L. subsp. trivialis und Poa trivialis subsp. sylvicola (GUSS.) H. LINDB. fil. Die "typische" Subspecies trägt keinen Autorennamen. Die hier als Beispiel gewählte Unterart sylvicola trägt in Klammern den Namen GUSS (= GUSSONE), und ohne Klammer H. LINDB. fil. (= H. LINDBERG filius). Ein in Klammern genannter Autor hat eine bestimmte Unterart (Art oder Gattung) als erster beschrieben, sie aber in eine andere Rangstufe der gleichen oder einer anderen Art (oder Gattung) gestellt. Der nicht in Klammern stehende Autor hat die hier präsentierte Rangstufe festgelegt. Familiennamen ergeben sich aus dem Namen der für die Familie typischen Gattung. Daher gilt z.B. für die Gräser der Name Poaceae. Für einige wenige Familien bleiben zusätzlich traditionelle Namen gültig, so können die Gräser auch heute noch als Gramineae bezeichnet werden. Ferner sind noch die folgenden Ausnahmen zulässig: Palmae, Cruciferae, Leguminosae, Guttiferae, Umbelliferae, Labiatae und Compositae.
Ordnungsnamen leiten sich vom Namen der für die Ordnung typischen Familie ab, die Endung lautet ...ales (z.B. Poales). Die Namen der Klassen und Unterklassen werden in ähnlicher Weise gebildet. Ihre Endungen sollten sein: Bei den Algen .....phyceae (Klassen) und . ...phycidae (Unterklassen). Bei den Kormophyten (Gefäßpflanzen):.....atae oder ...opsida (Klassen) und ....idae (Unterklassen). Auch hier, vor allem bei den Blütenpflanzen, dürfen konventionelle Namen wie Angiospermae, Dikotyledonae u.a. weiter verwendet werden.
Die den einzelnen Taxa zugeordnete Rangstufe ist verschieden in den Systemen verschiedener Autoren, da sich die Rangstufen oberhalb der Art kaum absolut definieren lassen; vielmehr ist primär ihre relative Position in der Rangstufenhierarchie wichtig. So werden z.B. die Angiospermen traditionellerweise als Unterabteilung "Magnoliophytina" der Abteilung Spermatophyta bewertet. Bei CRONQUIST et al. (1966) etwa haben sie dagegen den Rang der Abteilung. Systematisch viel wesentlicher als die absolute Rangstufe ist jedoch die relative Position der Angiospermen zu den Gymnospermen, bzw. zu den Cycadophytina und den Coniferophytina.
Eine Klassifikation von Pflanzen nach einem hierarchischen System ist geeignet, einzelne Arten und Merkmalsgruppen mit der EDV zu bearbeiten. Es sind Verfahren entwickelt worden, die unter der Bezeichnung Numerische Taxonomie bekanntgeworden sind. In je mehr Merkmalen Arten (oder andere Taxa) übereinstimmen, desto näher sollten sie untereinander verwandt sein. Je mehr Arten man miteinander vergleichen möchte, um so mehr Merkmale (Merkmalsausprägungen) muß man dabei heranziehen. Numerische Taxonomie ist ein interessanter Ansatz, zu dem bereits Ende des vorigen Jahrhunderts erste Vorstellungen entwickelt wurden. In der ersten Hälfte dieses Jahrhunderts wurden sie erfolgreich und in umfangreichem Maße von A. H. STURTEVANT an Drosophila-Arten erprobt. Ein Versuch, sie zur Klassifikation von Pflanzen anzuwenden, wurde in den fünfziger Jahren unternommen (R. R. SOKOL P. H. A. SNEATH, 1962). Er hat sich nicht durchsetzen können. Schwachpunkte liegen in der Tatsache, daß nicht alle Merkmale das gleiche Gewicht haben, daß weiter jedes Merkmal einen adaptiven Wert hat, die Selektion je nach Ort und Zeit unterschiedlich wirkt und dadurch eine Variation der Merkmale zutage tritt, die nur schwer standardisierbar ist. Vor allem aber auch, weil erwiesen ist, daß die Zahl unterschiedlicher Merkmale für eine Artabgrenzung belanglos ist.
Klassifikationen beruhen immer auf einer Bewertung von Merkmalen. Diese sagen aber a priori nichts über ihren adaptiven Wert oder über phylogenetische Zusammenhänge aus. So hat z.B. THEOPHRAST die Blütenpflanzen in Holzpflanzen und Kräuter unterteilt, und LINNÉ schlug vor, sie nach der Zahl der Staubblätter zu gruppieren. Beide Einteilungen gelten als künstlich. Seit Anfang des vergangenen Jahrhunderts ist man bemüht, sie durch ein natürliches System zu ersetzen, aber erst seit DARWIN kam das Bestreben hinzu, hierin die phylogenetische Verwandtschaft der Arten und anderer Taxa zum Ausdruck zu bringen. Das Ziel solcher Überlegungen wäre die Erstellung eines Stammbaums der Pflanzen. Einen ersten Versuch hierzu unternahm E. HAECKEL.
Eine vollständige Darstellung müßte die genaue Position einer jeden Art wiedergeben. Das ginge aber nur, wenn man die gemeinsamen Vorfahren rezenter und fossiler Arten kennen würde und wüßte, wie sie sich weiterentwickelt haben. Im einfachsten Fall wäre ein Stammbaum dann durch ein Schema aufeinanderfolgender dichotomer Verzweigungen darzustellen. Stammbäume dieser Art mit z.T. hoher Präzision kennen wir von einigen Stämmen des Tierreichs, bei Pflanzen ist man jedoch über einige mosaikartige Ansätze nicht hinausgekommen.
Es gibt zwar unangreifbare Beweise für die Entstehung der wichtigsten Abteilungen, Klassen und Unterklassen, oder für die der Arten in einigen Gattungen, doch sind wir von einem vollständigen Bild weit entfernt. Hierfür gibt es mehrere Gründe:
Es gibt wenige fossile Pflanzenreste, die für die Rekonstruktion der Abstammung einzelner Taxa brauchbar wären. Es gibt Versteinerungen, aus denen man Dünnschliffe herstellen kann, die guten mikroskopischen Präparaten in nichts nachstehen. Mit Dokumenten dieser Art sowie durch Auswertung von Abdrücken pflanzlicher Oberflächenstrukturen, konnte z.B. die Entwicklung der ersten Landpflanzen nachvollzogen werden. Doch fossile Urkunden eignen sich - von wenigen Ausnahmen abgesehen - kaum, um die Entstehung von Arten aus gemeinsamen Vorfahren erkennen zu können.
Die Ergebnisse der Evolutionsforschung zeigen, daß die Evolution von Pflanzen, vor allem die der Angiospermen, nicht nur auf divergierende, sondern auch auf konvergierende Entwicklungen zurückzuführen ist.
Das biologische Artkonzept veranschaulicht, daß sich eine Art unterschiedlich weiterentwickeln kann. Durch adaptive Radiation können Teilpopulationen verschiedenartige neue Lebensräume erschließen, und jede von ihnen kann sich zu einer neuen Art entwickeln. Aus einer Art können demnach nicht nur zwei, sondern n neue Arten hervorgehen, deren verwandtschaftliche Abstände zur Ausgangsart gleich sein können. Artbildung erfolgt auf diese Weise allerdings nur selten gleichzeitig, denn die Erschließung neuer Lebensräume erfolgt meist nacheinander. Wir haben aber nur selten die Möglichkeit, den zeitlichen Ablauf zu ermitteln oder zu rekonstruieren. Eine Pflanze produziert oft Hunderte oder mehr Samen, deren Genotypen sich stark voneinander unterscheiden können. Wenn die Umweltbedingungen es zulassen (oder fordern), können gleichzeitig verschiedene Genotypen selektiert werden, die sich ggf. zu neuen Arten weiterentwickeln können. Sie alle entstammen nur einem Individuum (einer Art). Selbst wenn dieser Extremfall als zu theoretisch abgetan werden sollte, bleibt er ein gutes Beispiel, um die Fragwürdigkeit eines Systems mit nur dichotomen Verzweigungen zu veranschaulichen.
Die vorgetragenen Schwierigkeiten stehen der Rekonstruktion der Abstammungsverhältnisse einzelner Arten (Gattungen, Familien, usw.) entgegen. R. DAHLGREN (Botanisches Museum, Universität Kopenhagen) schlug daher (1975, 1977) vor, statt eines Längsschnitts durch einen "Stammbaum" einen Querschnitt durch seine Krone zu legen, aus dem die verwandtschaftlichen Beziehungen rezenter Arten zueinander ablesbar sind. Hier werden die phylogenetischen Zusammenhänge durch die gegenseitige Anordnung der Gruppen zueinander symbolisiert. Sie spiegeln nicht nur den unterschiedlichen Umfang einzelner Taxa wider, sondern zeigen zugleich, wie komplex die Beziehungen der Gruppen untereinander sind und welche Affinitäten zwischen ihnen bestehen.
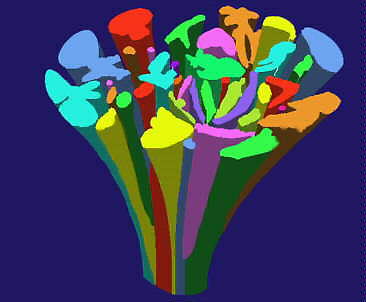
Schema eines "imaginären" Stammbaums der Angiospermen. Der Begriff "Strauch" wäre passender. Der derzeitige Zustand ist durch einen Querschnitt wiedergegeben. (Nach R. DAHLGREN, 1980)
|
|