Die Evolution der Pilze hängt weitgehend von der Weiterentwicklung und Ausbreitäng grüner Pflanzen ab. Die Mehrzahl der Pilzarten lebt saprophytisch, einige wenige parasitisch. Diese sind, zumindest in bestimmten Phasen ihres Lebenszyklus, auf ein Wirkstoffangebot (z.B. ein Angebot an Vitaminen) angewiesen, wie es in dieser Form nur von lebenden Zellen bereitgestellt werden kann. Es ist auffallend, daß Pflanzenkrankheiten in der Natur relativ selten sind. Die Ursachen dafür sind wirkungsvolle Abwehrmechanismen, die einer Ausbreitung parasitärer Pilze entgegenstehen. Dabei wäre in erster Linie die feste Zellwand, einschließlich aller Ein- und Auflagerungen (z.B. Kutikula), zu nennen, die ein Eindringen von Pilzen, Bakterien, Viren u.a. ins Gewebe und in die Zellumina verhindert. Zum anderen sei auf das weite Spektrum an sekundären Pflanzenstoffen verwiesen, von denen viele fungizid und/oder bakterizid sind. Oft werden derartige Substanzen erst nach einer Induktion, d.h., erst nach einer Infektion produziert (z.B. die Phytoalexine).
Ferner ist bemerkenswert, daß pflanzliche Parasiten (das gilt gleichermaßen für Pilze, Bakterien und Viren) streng wirtsspezifisch sind. Einige von ihnen sind auf einen Wirtswechsel angewiesen, durch den sich das eine Entwicklungsstadium des Parasiten in einem, das darauffolgende in einem anderen (mit dem ersten phylogenetisch nicht nah verwandten) abspielt. Wegen der hohen Wirtspezifität können Parasiten in Monokulturen (Landwirtschaft) weit mehr Schäden anrichten als in artenreichen Pflanzengesellschaften. Der jährliche Verlust an Nahrungsmitteln entspricht einer Menge, die zur Versorgung von 300 Millionen Menschen ausreichen würde.
Parasitäre Pilze verfügen über mindestens drei Strategien, um an pflanzliche Inhaltsstoffe zu gelangen:
Sie produzieren zellwand- und kutikulaabbauende Enzyme und .
Sie produzieren Toxine, die die Aktivität von Wirtszellen herabsetzten, ggf. sogar vollständig inhibierten.
Sie produzieren pflanzeneigene Substanzen, z.B. Hormone, und greifen damit in das hormonelle Gleichgewicht in der Pflanzenzelle ein, was Störungen im Wachstums- und Differenzierungsprozeß der Zellen und Gewebe zur Folge hat. Ein Beispiel: Gibberella fujikuroi sondert Gibberelline ab, die das Wachstum der Wirtspflanzen (hier Reis) beeinflussen. Die seinerzeit durchgeführten Untersuchungen führten zur Entdeckung der Phytohormonklasse der Gibberelline.
Es gibt eine sehr umfangreiche Literatur über parasitäre Pilze. Viele der Arbeiten sind aus wirtschaftlichen Erwägungen heraus entstanden. Die Mehrzahl befaßt sich mit der Klassifikation der Pilze, ihren Lebenszyklen, den Krankheitssymptomen der Pflanzen und deren Diagnostik, dem Wirtsspektrum, und der Suche nach Resistenzfaktoren der Wirte. Hingegen ist der molekulare Wirkungsmechanismus der Reaktion von Pflanzen nach Pilzinfektion bislang nur an wenigen Beispielen geklärt worden und wegen der Vielfalt der Möglichkeiten können diese Fälle nur bedingt verallgemeinert'werden.
Die Resistenz der Wirte beruht - außer auf den eingangs skizzierten allgemeinen (unspezifischen) Abwehrmechanismen- auf Bildung spezifischer (gegen bestimmte Pilze gerichteter), genetisch determinierter Produkte. Genetische Analysen ergaben, daß die Resistenz einmal auf dem Vorhandensein dominanter Allele der entsprechenden Gene beruht und daß es zum anderen unabhängig voneinander vererbbare Resistenzgene gibt.
Man unterscheidet zwischen zwei Formen des Parasitismus:
Nekrotropher Parasitismus: Die Infektion führt zur Gewebezerstörung und damit zum Tod der Pflanze. Die Pilze sind meist nur fakultativ parasitär; sie können sich ebenso gut saprophytisch in totem oder absterbendem Pflanzenmaterial vermehren. |
|
Biotropher Parasitismus: Hier leben Parasit und Wirt, zumindest über längere Zeiträume hinweg, zusammen. Der Parasit entnimmt dem Wirt Nähr- und Wuchsstoffe, tötet ihn jedoch nicht ab. Die meisten biotrophen Pilze sind obligate Parasiten. Eine saprophytische Phase können sie nur bedingt überstehen; vor allem die Fruchtkörperbildung ist an das Vorhandensein des Wirts gebunden. Nur in Ausnahmefällen gelang es, einzelne (vegetative) Stadien dieser Pilze in zellfreien Nährmedien zu kultivieren. |
Parasitäre Pilze findet man in allen Pilzklassen, Wirte in allen systematischen Gruppen der Pflanzen (und der Blaualgen). Das Reich der Pilze gliedert sich in die Myxomycota oder Schleimpilze und die Eumycota, die eigentlichen Pilze.
Einer der wirtschaftlich wichtigsten Vertreter der parasitären Myxomycota ist Plasmodiophora brassicae, der Erreger der Kohlhernie, einer Krankheit, deren Symptome sich im Wurzelbereich zahlreicher Cruciferen-Arten manifestieren. Das veränderte Differenzierungsmuster des infizierten Wirtsgewebes beruht u.a. auf einer Zunahme des Auxingehalts um das 50-100 fache, einer Cytokininzunahme um das 10-100fache und (als Folge davon?) einer Erhöhung des Ploidiegrads in den Kernen jener Gewebezellen (I. C. TOMMERUP und D. S. INGRAM, 1971).
Spongospora subterraneae der Erreger des Pulverschorfs der Kartoffel, kann als Vektor für das Kartoffel-mop-top-Virus dienen.



Myxostelida - Sporen (Ruhestadien), Myxamöben entlassend - Trichiales - junge Sporangien von Trichia decipiens - Trichiales - Plasmodiocarp (vielkerniges Plasmodium) von Hemitrichia serpula
© Bryce KENDRICK
Eumycota. Phytophthora-Arten sind u.a. auf Solanaceen als Wirtspflanzen angewiesen. Phytophthora infestans ist der Erreger der Kraut- und Knollenfäule der Kartoffel. Eine frühe Zerstörung des Laubs, und die dadurch bedingte Reduktion der Photosyntheserate, führt zu gravierenden Ernteeinbußen. Das Wirtsspektrum ist, im Gegensatz zu dem mancher anderer Phytophthora-Arten extrem eng (Kartoffel, Tomate und wenige andere). Der Pilz war Mitte des vorigen Jahrhunderts (1845/47) die Ursache einer großen Hungerkatastrophe in Irland; die Folge davon war eine starke Auswanderungswelle in die USA. Phytophthora cambivora ist für ein sich z. Zt. in Deutschland ausbreitendes Erlensterben verantwortlich (Wurzelhalsfäule der Schwarzerle).

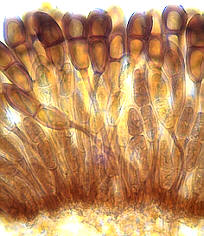
Es gibt vier Typen unterschiedlich aussehender Asci, wobei es zwei Grundtypen gibt:
unitunicat und bitunicat.
Die Abbildung zeigt den bitunicaten Typ.
|
Zu den Taphrina-Arten gehören auch jene, die die sogenannten Hexenbesen auf zahlreichen Wirtsbäumen hervorrufen. Diese Erscheinung geht auf eine Erhöhung der Zahl der Vegetationspunkte zurück, wodurch ein irreguläres büschelförmiges Verzweigungsmuster der befallenen Äste sichtbar wird.
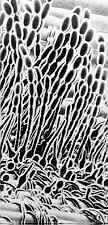 Die Ordnung Erysiphales enthält die Familie Erysiphaceae (Echter oder Weißer Mehltau).
Der Echte Mehltau ist ein Sammelname für eine
Vielzahl von Erysiphaceen-Arten, die auf zahlreichen Angiospermen
parasitieren: Erysiphe graminis (Mehltau auf Gräsern),
E. communis (auf Kürbis), E. polygoni
(auf Erbsen, Klee u.a. Leguminosen). Das Pilzmycel breitet sich
in der Regel auf der Blattoberseite (oder der Unterseite) aus,
lediglich in die Epidermiszellen werden vereinzelt Haustorien
eingesenkt.
Die Ordnung Erysiphales enthält die Familie Erysiphaceae (Echter oder Weißer Mehltau).
Der Echte Mehltau ist ein Sammelname für eine
Vielzahl von Erysiphaceen-Arten, die auf zahlreichen Angiospermen
parasitieren: Erysiphe graminis (Mehltau auf Gräsern),
E. communis (auf Kürbis), E. polygoni
(auf Erbsen, Klee u.a. Leguminosen). Das Pilzmycel breitet sich
in der Regel auf der Blattoberseite (oder der Unterseite) aus,
lediglich in die Epidermiszellen werden vereinzelt Haustorien
eingesenkt.
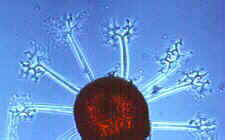
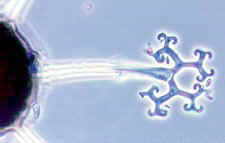
Erysiphales - Rasterelektronenmikroskopische Aufnahme von Konidien von Erysiphe auf dem Blatt einer Wirtspflanze (Bild links) - dichotom verzweigte akrosomale Fortsätze von Microsphaera - Asci von Microsphaera, mit dichotom verzweigten Fortsätzen
© Bryce KENDRICK
Von Erysiphe graminis sind zahlreiche Unterarten (Rassen, Varietäten?) auf Getreidearten spezialisiert, einzelne beispielsweise wachsen auf Weizen, aber nicht auf Gerste; für andere gilt genau das Umgekehrte.
Ceratocystis ulmi und verwandte Arten sind Verursacher des Ulmensterbens. 1927 wurde die Krankheit in England festgestellt, zwischen 1930 und 1940 waren dort etwa 10 Prozent aller Ulmen befallen. Im trockenen Sommer 1947 breitete sie sich epidemisch auch über Deutschland aus.
Botrytis cinerea (Grauschimmel) ist ein wenig spezialisierter Parasit; er befällt bei feuchter Witterung Salatblätter ebenso wie eine Vielzahl saftiger Früchte (Tomaten, Erdbeeren usw.).
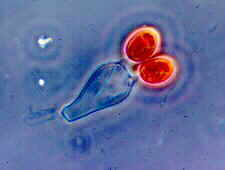
Basidium eines Basidiomyceten, Gastrocybe, mit symmetrischer Anordnung der Basidiosporen.
|

UREDINALES: Teleutosporenlager. Die gestielten zweizelligen Teleutosporen entstehen nach Wirtswechsel (z.B. bei Puccinia) im Spätsommer auf Getreidepflanzen und überwintern am Boden. Aus diesen Probasidien entwickeln sich die eigentlichen, septierten Basidien (Aufn.: W. KASPRIK).
Die Voraussetzung dafür sind jedoch milde Winter, denn die dikaryotischen asexuellen Uredosporen ("Sommersporen") können lange, kalte Winter nicht ertragen. Andererseits können Pilzsporen durch Wind über Tausende von Kilometern verbreitet werden, so daß eine lokale Ausrottung der Berberitze keinerlei dauerhaften Schutz vor Puccinia bietet. Auch diese Art besteht aus einer Reihe von Unterarten, von denen eine auf Weizen, eine andere auf Hafer, eine dritte auf Roggen usw. spezialisiert ist. Puccinia graminis gehört zu den wenigen parasitären Pilzen, die zellfrei auf Agar (mit Zusatz von Hefeextrakt) gehalten werden können.

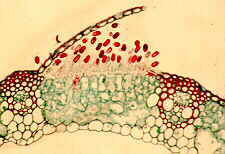
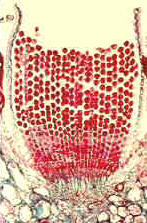
Uredinales - Uredosporen von Puccinia graminis tritici auf der Epidermis eines Weizenblattes (Triticum). - Querschnitt durch einen Sorus mit Uredosporen von Puccinia graminis tritici in einem Weizenblatt. - Querschnitt durch ein Aecidium von Puccinia graminis tritici.
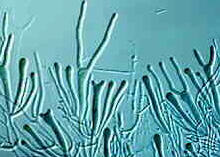
Dacrymycetales - Basidien, aussehend wie Stimmgabeln von Dacrymyces in verschiedenen Entwicklungsstadien
© Bryce KENDRICK
Fungi imperfecti. Dies ist ein Sammelbegriff für Pilzarten, die keine Fruchtkörper ausbilden und die deshalb auch recht schwer klassifizierbar waren, aufgrund ultrastruktureller Untersuchungen nimmt man heutzutage an, sie (die meisten?) sind den Askomyceten zuzuordnen. Hierher gehören einige Arten, die in den letzten Jahren als Versuchsobjekte zum Studium der molekularen Vorgänge bei der Infektion von Pflanzen benutzt wurden. Wir werden uns daher im übernächsten Abschnitt ausführlich mit ihnen auseinandersetzen müssen.
Parasitäre Wasserpilze. Da parasitäre Wasserpilze wirtschaftlich unbedeutend sind, sind sie relativ unbekannt. Es gibt nur wenige Wissenschaftler, die sich mit ihnen intensiv auseinandergesetzt haben.
Über 200 filamentöse marine Pilze wurden beschrieben, von denen etwa ein Viertel bis ein Drittel auf Algen parasitiert. Die Mehrzahl der Pilze gehört zu den Askomyceten. Als Wirte kommen Braun-, Rot- und Grünalgen sowie Diatomeen in Betracht. bes an Lagenisma coscinodisca, einem Diatomeenparasiten, elektronenmikroskopisch dokumentiert,
Die Parasiten der Süßwasseralgen gehören vornehmlich zu den Chytridien, die 1856 von A. BRAUN (Berlin) entdeckt wurden, und die er wie folgt beschreibt:
"Das ganze Pflänzchen besteht aus einer einfachen blasenartigen Zelle, welche oft mit einer wurzelartigen Verlängerung in die Zellen des Nährorganismus eindringt."
Chytridien bilden keine Hyphen aus, statt dessen wird ein (nichtzelluläres), teilweise recht umfangreiches Rhizoidsystem gebildet, mit dem die Wirtszellen erfaßt, z.T. auch umschlossen werden. Für nahezu jede der bekannten Algenarten konnte ein für sie spezifischer Parasit gefunden werden. Der Pilzbefall sollte nicht unterschätzt werden; in vielen Fällen konnte gegen Ende einer "Wasserblüte" (dem massenhaften Auftreten einer Algenart) eine Infektion nahezu aller verbliebenen Zellkolonien festgestellt werden.
Chytridien befallen sowohl Blaualgen (Cyanophyta) als auch Volvocales und Chlorococcales. Da sich die drei genannten Algengruppen u.a. durch die chemische Zusammensetzung ihrer Zellwände voneinander unterscheiden, müssen die entsprechenden Parasiten über die verschiedensten wandabbauenden Enzymsysteme verfügen. Die Zerstörung einer Zelle erfolgt nur nach einem Zell-Zell-Kontakt. Das heißt, daß der Pilz kein lytisches Agens ins umgebende Medium sezerniert, um somit ggf. sämtliche Zellen einer von Gallerte umgebenen Kolonie zu zerstören.
Jahrelange, populationsdynamische Untersuchungen (in Seen des südenglischen Lake Districts) an Diatomeen- und Cyanophyceenparasiten ergaben, daß Pilzbefall zu Beginn eines Populationswachstums die Individuenzahl drastisch reduzieren kann sowie den nachfolgenden Niedergang der Population nachhaltig beschleunigt. Die Reduktion der Individuenzahl einer Algenart wird aber in der Regel durch einen Anstieg der Populationsgröße einer anderen Art kompensiert (H. M. CANTER; Freshwater Biological Association, Ambleside). Derartige quantitative Analysen - und eine größere Beachtung der Konsequenzen des Pilzbefalls sind mögliche Erklärungen für den von Algologen häufig ermittelten Sachverhalt, daß eine Algenart in einem Jahr dominierend ist, im darauffolgenden gar nicht oder nur wenig in Erscheinung tritt.
Der Befall von Algenkolonien durch Chytridien ist jedoch keineswegs stets der alleinige Grund für den Zerfall einer Wasserblüte; andere Gründe, z.B. Nährstoffmangel (Phosphatmangel) oder witterungsbedingte Änderungen, sind oft ausschlaggebend, vor allem dann, wenn sich der Pilzbefall in Grenzen hält und nur wenige Zellen einer Kolonie (z.B. bei der Blaualge Micocystis aeruginosa) zerstört werden.
|
|