Seit langem war man bemüht, pflanzliche Gewebe auf künstlichem Nährmedium zu kultivieren. Die ersten Ansätze hierzu gehen auf den österreichischen Pflanzenanatom G. HABERLANDT (1854-1945, Professor in Graz, später in Berlin) zurück. Große Probleme bereitete anfangs die Zusammensetzung eines geeigneten Nährmediums; erst als der holländische Pflanzenphysiologe J. v. OVERBEEK 1941 entdeckte, daß ein Zusatz von Kokosnußmilch zum Medium das Wachstum von Pflanzenembryonen und Gewebekulturen drastisch steigerte, wurde der eigentliche Durchbruch erzielt. Kokosnußmilch ist ein Produkt des Endosperms, die auch unter natürlichen Bedingungen wachstumsfördernd auf den sich entwickelnden Kokosnußembryo wirkt. Damit war natürlich sofort die Frage aufgeworfen, worauf die Wachstumsförderung beruhe und welche Komponenten dafür verantwortlich zu machen seien.
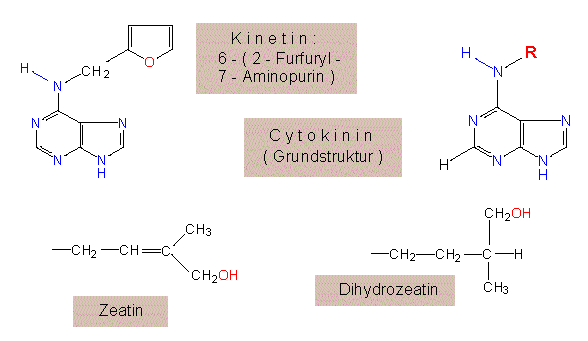
Im Gegensatz zu einer Auxinwirkung wird nicht das Streckungs-, sondern das Teilungswachstum von Geweben gefördert. 1955 fanden C. O. MILLER und F. SKOOG von der University of Wisconsin in Madison, daß gealterte oder autoklavierte DNS-Präparate ebenso wirksam sind, frische DNS-Präparate hingegen nichts bewirken. Als wirksame Komponente wurde schließlich das Adeninderivat 6-Furfurylaminopurin (= Kinetin) identifiziert.
Kinetin ist physiologisch außerordentlich aktiv, obwohl gerade diese Substanz aus keiner Pflanzenzelle isoliert werden konnte. Statt dessen fand man ein weites Spektrum ihm ähnlicher Verbindungen. Die erste aus einer natürlichen Quelle (unreifen Maiskörnern) isolierte Komponente war das Zeatin (D. S. LETHAM et al., 1964).
Die Formeln dieser Verbindungen machen es wahrscheinlich, daß Cytokinine (= Sammelname für Verbindungen dieses Typs) in den Nukleinsäurestoffwechsel eingreifen. Doch sollte man diese Vermu4ung nicht unkritisch hinnehmen. Aus tierischen Zellen ist nämlich das cAMP als Second messenger bekannt. Obwohl es sich vom ATP herleitet und selbst den Nukleotiden zuzurechnen ist, hat es in tierischen Zellen (und Mikroorganismen) wenig mit der Regulation des Nukleinsäurestoffwechsels zu tun. Vielmehr sind zahlreiche Proteine identifiziert worden, mit denen es in Wechselwirkung tritt. Deren Konformationen und Aktivitäten werden nach cAMP-Bindung drastisch verändert.
Zurück zu den Cytokininen. Es sind in verschiedenen Laboratorien mittlerweile Hunderte verschiedener Cytokininderivate synthetisiert worden. Viele von ihnen sind ebenso wirksam wie das Kinetin. Für biologische Wirksamkeit ist die Substitution des N-Atoms 6 (im 6er-Ring) erforderlich. Jeder Austausch eines Ringatoms durch ein anderes führt zu einem Aktivitätsverlust. Eine Alkylgruppe als Substituent am N6 hat die höchste Wirkung bei Kettenlängen von 5 C-Atomen. Eine Doppelbindung und/oder eine Hydroxylgruppe steigern die Aktivität um ein Mehrfaches. Es sieht also ganz so aus, als würden alle Modifikationen, die zu mehr planaren Strukturen führen, die Aktivität erhöhen. Damit wäre u.a. auch erklärt, weshalb der Furfurylring im Kinetin so außergewöhnlich wirkungsvoll ist. Zu den natürlich vorkommenden, sehr aktiven Cytokininen gehört das IPA (Isopentenyladenin) aus Gewebekulturen des Tabaks. In den Zellen entsteht es durch Anknüpfung der Seitenkette an einen in tRNS (tRNSser und tRNStyr) inkorporierten Adeninrest.
Wie die Auxine können Cytokinine glykosyliert oder an Aminosäuren oder Proteine geknüpft sein, und damit - zumindest vorübergehend - in einen inaktiven Zustand überführt werden.
Biologische Aktivitäten. Gewebekulturen, z.B. von Tabak oder Ahorn (Acer pseudoplatanus), können nur nach Cytokininzugabe gedeihen. Cytokinine steigern neben der DNS-Replikationsrate die allgemeine RNS- und die Proteinsyntheserate, verlangsamen Alterungserscheinungen (Seneszenz) und stimulieren die Dunkelkeimung lichtbedürftiger Samen. Daneben wurde eine Reihe selektiver Wirkungen beobachtet wie:
| Induktion von Isocitrat-Lyase und Protease-Aktivität in abgetrennten Kürbis-Kotyledonen | |
| Aufhebung des Thiamin-Bedarfs wachsender Kalluskulturen des Tabaks durch Induktion einer Thiaminsynthese. | |
| Förderung der Auxinsynthese in Tabakgewebekulturen. | |
| Steigerung der Aktivität der Carboxydismutase und NADP-Glycerinaldehydphosphatdehydrogenase in etiolierten Reiskeimlingen. | |
| Förderung von Knospenentwicklung sowie der Keimung einiger Samen und Förderung der Akkumulation von Nitratreduktase in einigen Embryonen. |
Cytokinine werden in der Regel in der Wurzel, in jungen Früchten und in Samen gebildet. Über das Xylem wandern sie in Sproßorgane ein. Bei unterbrochenem Nachschub, z.B. in abgeschnittenen Sprossen, altern diese schneller als wurzeltragende Sprosse. Durch Kinetinzusatz kann dem Alterungsprozeß Einhalt geboten werden. Bildung von Adventivwurzeln - und damit erneute Versorgung mit Cytokininen - stellt den alten Zustand wieder her.
Das Zusammenwirken von IES und Kinetin und die relativen Verhältnisse zueinander entscheiden darüber, ob sich z.B. ein Tabakkallusgewebe zu Wurzeln und/oder zu Sprossen differenziert. Offensichtlich stehen einer undifferenzierten Zelle zwei Wege zur Weiterentwicklung offen: Entweder sie vergrößert sich, teilt sich, vergrößert sich usw., oder sie wächst (ohne Teilung) in die Länge (Zellstreckung). Eine sich häufig teilende Zelle bleibt weitgehend undifferenziert, während gestreckte Zellen eine Tendenz zur Differenzierung und damit zu einer Spezialisierung erwerben. Wir wissen, daß IES alleine die Zellstreckung fördert, während Kinetin alleine nichts bewirkt. Beide zusammen fördern schnelle Zellteilungen.
Bei einem Verhältnis IES/Kinetin von 3 mg/0,2 mg/l wächst ein Kallusgewebe (es finden Zellteilungen statt); wird der Kinetinwert auf 0,02 mg/l gesenkt, wird eine Wurzelbildung induziert und wird das Verhältnis zugunsten des Kinetins verschoben (0,03 mg/1,0 mg/l), entwickeln sich Sprosse.
Das IES/Kinetin-Verhältnis reguliert u.a. auch das Lignin-Pektin-Verhältnis in Zellkulturen des Tabaks. Bei hohen Kinetin- und niedrigen IES-Mengen ist der Ligninanteil im Verhältnis zum Pektinanteil hoch, bei umgekehrten Verhältnissen erhält man dementsprechend reziproke Werte.
Die Wechselwirkung zwischen IES und den Cytokininen ist Calcium-abhängig. Durch Zusatz von Calciumionen wird das Wachstumsmuster von Zellstreckung zu Zellteilung verschoben. Hohe Calcium-Konzentrationen unterbinden die Streckung der Zellwand; es werden keine neuen Strukturelemente mehr eingelagert.
Wie schon am Beispiel der Auxine erläutert, gibt es auch für Cytokinine keinen einzigartigen Wirkungsmechanismus. Die Vielfalt der hormonell induzierten Erscheinungen weist auf die Existenz unterschiedlicher Rezeptoren hin.
|
|