Worauf beruht die hochgradige Spezifität der Wirt-Parasit-Beziehungen? Um diese Frage zu beantworten, ist es zweckmäßig, sie in eine Reihe von Detailfragen zu untergliedern und jeden Teilaspekt getrennt zu behandeln:
![]() Wie gelangt ein Pilz ins pflanzliche Gewebe?
Wie gelangt ein Pilz ins pflanzliche Gewebe?
![]() Wie überwindet er das Hindernis Zellwand?
Wie überwindet er das Hindernis Zellwand?
![]() Welchen Einfluß haben Toxine auf die Plasmamembran?
Welchen Einfluß haben Toxine auf die Plasmamembran?
![]() Wie reagieren Pflanzenzellen auf die Aktivitäten der
Pilze?
Wie reagieren Pflanzenzellen auf die Aktivitäten der
Pilze?
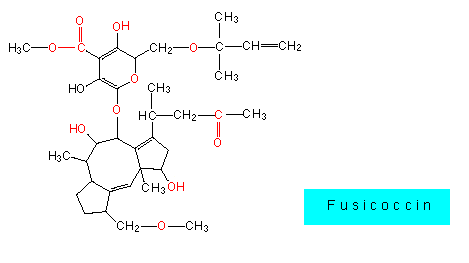
Im einfachsten Fall dringen auskeimende Sporen oder Pilzhyphen durch Wundstellen der Abschlußgewebe oder deren Auflagen (es genügt meist schon eine eingerissene Kutikula), oder durch offene Stomata in pflanzliche Gewebe ein. Einige Spezialisten, z.B. Fusicoccum amygdali (wie die meisten der nachfolgend erwähnten Beispiele ein Fungus imperfectus), sezernieren ein Terpenoid (Fusicoccin), das den Kalium-Influx in Stomatazellen erhöht und damit eine permanente Stomataöffnung induziert. Die Folge ist ein starker Wasserverlust der Pflanze, an dem sie letztlich zugrunde geht. Fusicoccin wird daher als ein Welketoxin bezeichnet.
Die Toxine (hier zyklische Peptide) von Helminthosporium maydis, dem Erreger einer Maiskrankheit (befällt vornehmlich Mais des Genotyps Texas male sterility cytoplasm - CMS), der 1970 in den USA eine der schlimmsten pilzverursachten Epidemien hervorgerufen hat, reagieren genau umgekehrt: Sie hemmen die lichtinduzierte Kalium-Aufnahme durch die Stomata. Damit werden die Transpiration und folglich auch die Photosyntheseaktivitäten herabgesetzt (C. J. ARNTZEN et al., 1973).
Die Methoden zur Klärung dieses Problems wurden in der Arbeitsgruppe von P. ALBERSHEIM (Complex Carbohydrate Research Center, Athens, Georgia) erarbeitet. Ihr Versuchssystem bestand aus einem Pilz (meist C/lletrotrichum lindemuthianum) und der Wirtspflanze Phaseolus vulgaris. Die ebenfalls in dem Labor erzielten Ergebnisse auf dem Gebiet der Chemie pflanzlicher Zellwände waren eine günstige Voraussetzung zur Bearbeitung der hier anstehenden Fragen.
Untersucht wurden die vom Pilz ausgeschiedenen zellwandabbauenden Enzyme; als Substrat dienten isolierte Wände aus Zellen des Hypokotyls der Wirtspflanze. Gefunden wurden Polygalacturonasen und mit ihnen verwandte Enzyme, so vor allem alpha-Galactosidase, beta-Galactosidase, beta-Xylosidase und alpha-Arabinosidase. Diese Enzyme waren in der Lage, Wände aus fünf Tage alten Hypokotylstücken restlos zu verdauen, während die aus 18 Tage alten Hypokotylstücken dem enzymatichen Angriff weitgehend standhielten. Es waren bereits Sekundärwände vorhanden, für deren Abbau andere Enzyme erforderlich sind.
Es mag auffallen, daß die hier aufgeführten Enzyme ausschließlich die Pektin- und Hemicellulosefraktion der Wand, nicht aber das Cellulosegerüst angreifen. Die durch enzymatischen Teilabbau entstehenden Fragmente werden Oligosaccharine genannt. Wie es sich später zeigte, spielen auch sie bei der Pilzabwehr eine wichtige Rolle. Die Grundstruktur der Zellen bleibt demnach erhalten, sie wird - wie wir bereits im vorletzten Abschnitt sahen, nur lokal perforiert, die Zelle bleibt am Leben. Etliche der saprophytisch lebenden Pilze produzieren hingegen große Mengen an Cellulasen und decken durch den Celluloseabbau den eigenen Kohlenstoffbedarf.
Die Synthese der alpha-Galactosidase durch Colletrotrichum ist induzierbar. Sie erfolgt nur in Anwesenheit von Wänden aus nichtresistenten Wirten. Die Induzierbarkeit weist darauf hin, daß ein von der Wirtszelle erzeugtes Signal benötigt wird. Andererseits ließ sich auch zeigen, daß die Zellwände spezifische Inhibitoren der alpha-Galactosidase enthalten. Der aus Phaseolus vulgaris isolierte Inhibitor inaktiviert das Enzym aus Colletrotrichum vierzigmal effektiver als das entsprechende Enzym aus Fusarium oxysporum (einem Parasiten der Tomate). Es beeinflußt nicht das Enzym aus Sclerotium rolfsii. Der Inhibitor ist ein Glykoprotein mit Lektineigenschaften, d.h., mit einer Affinität zu bestimmten Zuckerresten. Das wiederum heißt, daß die pilzlichen Polygalactosidasen ebenfalls Glykoproteine sind und daß deren Glykosylierungsmuster offensichtlich für die Spezifität der Wirt-Parasit-Interaktionen entscheidend ist.
Eine Anzahl mittlerweile getesteter Pilzarten unterscheidet sich im Enzymmuster quantitativ und qualitativ von Colletrotrichum. Fusarium oxysporum z.B. bildet nach Infektion von Tomaten als erstes Polygalacturonsäure degradierende Enzyme. Rhizostonia solani produziert in zwei aufeinanderfolgenden Phasen zunächst die o.g. Enzyme, später werden Phenoloxydasen gebildet, die es dem Pilz ermöglichen, Sekundärwände anzugreifen.
Phytophthora infestans sezerniert zwei Polygalacturonasen, vier Galactanasen und zwei Pektinesterasen (die jeweiligen Enzyme unterscheiden sich durch Substratspezifität und Umsatzraten). Die Polygalacturoasen setzen aus Kartoffelzellwänden weniger als 6 Prozent der Kohlenhydrate frei, die Galactanasen 23 Prozent (M. C. JARVIS et al., 1981).
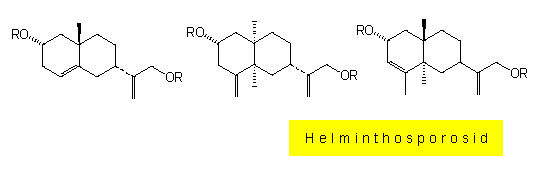
G. STROBEL und Mitarbeiter (Montana State University, Bozeman) befaßten sich mit Helminthosporium sacchari, dem Erreger der Augenfleckkrankheit des Zuckerrohrs. STROBEL stellte sich dabei weniger die Frage, warum eine Pflanze gegenüber einem Pilz resistent ist (das ist nämlich die Regel), sondern warum sie empfinglich ist. Pilzbefallene Zuckerrohrblätter enthalten das Mycel in kleinen, augenförmigen Infektionsherden, von denen aus sich mehrere zentimeterlange, rotbraun gefärbte Streifen parallel zur Blattachse ausbreiten. Da sie mycelfrei sind, lag es nahe, nach einem kleinen, diffusiblen Toxinmolekül zu suchen. Eine solche Substanz wurde aus infizierten Blättern isoliert und durch Kernspinresonanzanalyse und Massenspektroskopie zunächst als 2-Hydroxycyclopropyl-alpha-D-Galactopyranosid (Helminthosporosid) identifiziert, genauere Analysen zeigten jedoch, daß es sich um eine Mischung von drei isomeren Sesquiterpenalkaloiden handelt, die an beiden Molekülenden Digalactosylreste tragen. Diese liegen in der seltenen Furanoseform vor.
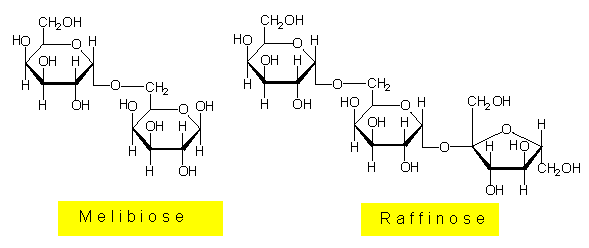
Helminthosporosid kommt in gesunden Blättern nicht vor. Besprüht man Zuckerrohrblätter mit einer Lösung dieses Toxins, erscheinen die pilz-spezifischen Symptome. Hierdurch angeregt, entwickelte G. W. STEINER (Hawaiian Sugar Planters Association) ein Schnellverfahren zur Selektion resistenter Zuckerrohrkeimlinge. Das Verfahren wird im Rahmen von Züchtungsprogrammen auf Hawaii routinemäßig eingesetzt. Das Helminthosporosid ähnelt alpha-Galactosiden, so dem Disaccharid Melibiose und dem Trisaccharid Raffinose. Beide Zucker kommen in pflanzlichen Geweben vor, und es ist bekannt, daß sie von Pflanzenzellen aktiv aufgenommen werden können.
Es zeigte sich, daß das Toxin an der Außenseite der Plasmamembran die gleichen Bindungsstellen wie diese Zucker besetzt, doch im Gegensatz zu ihnen nicht ins Zellinnere transportiert wird. Da die Zellen auf die alpha-Galactoside nicht angewiesen sind, konnte die Toxinbindung allein nicht die Ursache der Toxizität sein. Das alpha-galactosidbindende Protein ist ein Tetramer, bestehend aus vier gleichen Polypeptidketten des Molekulargewichts von je 12000 (= ca. 110 Aminosäuren). Es kommt auch in resistenten Zuckerrohrstämmen vor, doch bindet es dort weder die Zucker noch das Toxin. Nach milder Behandlung mit Detergentien ist es in den aktiven Zustand überführbar. Aminosäureanalysen ergaben, daß je nach Stamm ein bis vier der 110 Aminosäurereste verändert waren und daß dadurch die Molekülkonformation so verändert war, daß die Affinität zum Substrat verlorenging. In der Plasmamembran steht dieses Protein offenbar in Nachbarschaft zu einer Kalium-/Magnesium-ATPase, denn durch Toxinbindung wird deren Aktivität (allosterischer Effekt) erhöht. Die Kalium-Aufnahme steigt. Die Folgen sind erhöhte Wasseraufnahme, erhöhter osmotischer Druck, Schädigung der Membran und schließlich Lyse der Zelle.
Wie schon erwähnt, produzieren Pflanzenzellen ein weites Spektrum an sekundären Pflanzenstoffen, von denen viele ein Pilzwachstum unterbinden. Zusätzlich produzieren sie (alle?), als Reaktion auf eine Infektion, spezifische Phytoalexine (K. O. MÜLLER und H. BÖRGER, 1941).
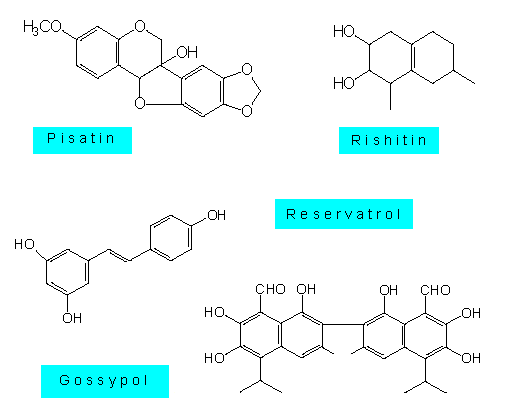
Das sind lipophile Substanzen, die von ihrer chemischen Struktur her den sekundären Pflanzenstoffen zuzuordnen sind. Die Phytoalexine der Solanaceen und Malvaceen sind meist Sesquiterpene, die der Leguminosen Isoflavonoide oder Polyacetylene, und die der Orchideen Dihydrophenanthrene. Manche Pflanzenarten, z.B. die Kartoffel, produzieren gleichzeitig mehrere, einander ähnliche Verbindungen. Über ihren Wirkungsmechanismus ist recht wenig bekannt. Manches deutet darauf hin, daß sie die Membraneigenschaften der Pilzzellen verändern; einige scheinen die oxydative Phosphorylierung zu blockieren, und von einigen anderen wiederum weiß man, daß sie DNS-Moleküle vernetzen können. Phytoalexine bieten keinen absoluten Schutz vor Pilzinfektionen. Sie richten sich vornehmlich gegen "nichtpathogene" Arten. Viele Parasiten sind in der Lage, sich vor ihnen zu schützen oder eigene Abwehrmechanismen aufzubauen.
Wodurch wird die Phytoalexinbildung induziert? In Zellkulturen der Sojabohne konnten P. ALBERSHEIM und Mitarbeiter Anfang der siebziger Jahre durch extern angebotene Poly- und Oligosaccharide eine Phytoalexinbildung hervorrufen. Als besonders wirkungsvoll erwies sich ein beta-l,3-Glucan mit seitlichen Verzweigungen (beta-3,6) und einem Molekulargewicht in der Größenordnung von 10000.
Solche Polysaccharide sind neben dem Chitin die Hauptstrukturkomponenten der Wand vieler Pilze (Asko- und Basidiomyceten). Das heißt nunmehr, daß ein Bestandteil der Pilzzellwand die Abwehrreaktion der Pflanze aktiviert. Bei langsamem Pilzwachstum können sich Phytoalexine anreichern und für den Pilz toxische Konzentrationen erreichen. Zeichnet er sich jedoch durch schnelles Wachstum aus, kann er sich ausbreiten (und die Pflanze schädigen), bevor die Phytoalexinabwehrreaktion zum Tragen kommt. Auslösende, spezifische Substanzen werden als "Elicitoren" (lat.: elegere = auslesen, auswählen) bezeichnet. Da es sich hier um Polysaccharide handelt, muß die Pflanze in ihrer Zellwand entsprechende Erkennungsmoleküle enthalten. Es sind per definitionem Lektine, und es sieht demnach so aus, als würde das Erkennen von Parasiten eine ihrer wichtigsten Funktionen sein. Das Lektin aus Weizenkeimlingen (WGA) hat eine hohe Affinität zu Chitin, und auch von ihm ist bekannt, daß es eine Ausbreitung von Pilzhyphen unterbindet.
Es fehlt aber noch ein Glied in der Ursache-Wirkung-Kausalkette. Man weiß noch nicht, wie das Signal (Bindung eines Polysaccharids an der Zellwandaußenseite) an deren Innenseite, und von dort durchs Plasmalemma hindurch, ins Zellinnere geleitet wird.
Hypersensitivität. Auf Induktion durch Pilze, Viren und andere Erreger, oder nach mechanischer Schädigung, können viele Pflanzenarten bzw. "resistente" Stämme, mit einer Hypersensitivitäts- oder Überempfindlichkeitsreaktion antworten. Sie ist durch das Absterben eines lokal begrenzten Gewebeabschnitts gekennzeichnet. Dadurch wird der Parasit von der Nährstoffzufuhr abgeschnitten, die Ausbreitungsmöglichkeit wird ihm genommen, und vielfach werden Wirkstoffe (phenolische Verbindungen) freigesetzt, die ebenfalls zu seinem Tod führen. Die toten, oft bräunlich gefärbten Bereiche der Pflanze nennt man Nekrosen.
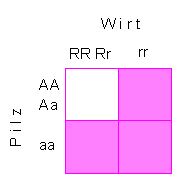 Hypersensitivität ist keine
als allgemein zu bezeichnende Reaktion, vielmehr ließ sich
zeigen, daß nur bestimmte Pflanzenarten auf bestimmte Parasiten
so reagieren. Genetische Analysen ergaben, daß sowohl im
Wirt als auch im Parasiten dominante Allele einzelner "Resistenzgene"
vorhanden sein müssen. Man spricht daher von einer Gen-zu-Gen-Wechselwirkung.
Die resistenzvermittelnden Gene werden mit R bezeichnet, die Gene
im Parasiten, die Avirulenz bewirken, d.h., keine Krankheitssymptome
hervorrufen, mit A. Die Kombination R - A entspricht damit einem
Stopp-Signal. Wirt und Parasit sind unter- und miteinander inkompatibel.
Diese auf den ersten Blick einfache Beziehung zeigt, daß
die Erkennung einer genetischen Analyse zugänglich ist und
daß die Wahrscheinlichkeit, durch züchterische Arbeit
zu resistenten Formen von (allen?) Kulturpflanzen zu gelangen,
recht hoch ist. Einschränkend sei jedoch betont, daß
es erstens eine ganze Anzahl voneinander unabhängiger R-Loci
gibt und daß auch im Pilz durch Mutation neue Formen entstehen,
die sich auf bislang resistenten oder durch Neuzüchtung resistent
gemachten Pflanzen ausbreiten können.
Hypersensitivität ist keine
als allgemein zu bezeichnende Reaktion, vielmehr ließ sich
zeigen, daß nur bestimmte Pflanzenarten auf bestimmte Parasiten
so reagieren. Genetische Analysen ergaben, daß sowohl im
Wirt als auch im Parasiten dominante Allele einzelner "Resistenzgene"
vorhanden sein müssen. Man spricht daher von einer Gen-zu-Gen-Wechselwirkung.
Die resistenzvermittelnden Gene werden mit R bezeichnet, die Gene
im Parasiten, die Avirulenz bewirken, d.h., keine Krankheitssymptome
hervorrufen, mit A. Die Kombination R - A entspricht damit einem
Stopp-Signal. Wirt und Parasit sind unter- und miteinander inkompatibel.
Diese auf den ersten Blick einfache Beziehung zeigt, daß
die Erkennung einer genetischen Analyse zugänglich ist und
daß die Wahrscheinlichkeit, durch züchterische Arbeit
zu resistenten Formen von (allen?) Kulturpflanzen zu gelangen,
recht hoch ist. Einschränkend sei jedoch betont, daß
es erstens eine ganze Anzahl voneinander unabhängiger R-Loci
gibt und daß auch im Pilz durch Mutation neue Formen entstehen,
die sich auf bislang resistenten oder durch Neuzüchtung resistent
gemachten Pflanzen ausbreiten können.
Heutzutage ist man auch bemüht mit Hilfe gentechnischer Verfahren Resistenz gegenüber Pilzen (und unabhängig davon gegenüber Viren) zu erzielen. Die Logistik und das Vorgehen eines solchen Experiments sind an der folgenden Stelle beschrieben:
|
|