

 |
Jede Zelle ist von einer Membran umgeben, und die meisten Zellen verfügen über umfangreiche intrazelluläre Membransysteme. Membranen grenzen das Zellinnere von der Umgebung ab, sie lassen Wasser, bestimmte lonen und Substrate in die Zelle hinein, sind für den Energiehaushalt der Zelle unentbehrlich und sondern Stoffe (Sekretions- und Exkretionsprodukte) ab. Sie bieten der Zelle Schutz und dienen der Abgrenzung des inneren Milieus von der Umwelt. Ohne Membranen würden Zellinhalte zerfließen, informationstragende Moleküle würden durch Diffusion verlorengehen, Stoffwechselwege einem thermischen Gleichgewicht zustreben, und das bedeutet bekanntlich den Tod eines lebenden Systems. |
Bild oben links: Elektronenmikroskopisches Bild einer Erythrozytenmembran (Plasmamembran, Plasmalemma). Die Membran erscheint als Doppelschicht, weil elektronendichtes Material zur Kontrastierung eingesetzt wird, das sich sowohl an der Innenseite als auch an der Außenseite der Membran ablagert. Die eigentliche Membran ist der Bereich zwischen den im Bild dunklen Schichten (J. D. ROBERTSON, Duke University, Durham)
Intrazelluläre Membranen trennen Kompartimente voneinander. Einige umschließen Vakuolen, andere Mitochondrien, Chloroplasten und den Zellkern. Kompartimente stellen ihrerseits klar voneinander getrennte Reaktionsräume dar. Alle biologischen Membranen besitzen in Organisation und Zusammensetzung eine Reihe von Gemeinsamkeiten:
Ihre Dicke beträgt durchschnittlich 70 Å.
Sie bestehen vornehmlich aus Lipiden und Proteinen.
Sie stellen eine Permeabilitätsschranke dar, und sie sind selektiv in bezug auf Substanzen, die hindurchgelassen werden.
Membranfragmente schließen sich stets zu Vesikeln.
Phospholipide stellen die in Membranen vorherrschende Molekülklasse dar. Sie sind amphipathisch, d.h., das Molekül besteht aus zwei Teilen, einem hydrophilen (polaren) Kopf (Phosphatgruppe und daran hängendem Rest R) und einem hydrophoben (lipophilen) Schwanz (Fettsäureanteil). Aufgrund dieser Struktur haben die Moleküle die Tendenz, sich zu ordnen und monomolekulare Schichten auszubilden, deren Ordnungsgrad von den Längen der Fettsäurereste und der Zahl der Doppelbindungen abhängt. Die Holländer E. GORTER und F. GRENDEL erkannten 1925, daß zwei derartige Schichten, in entgegengesetzter Orientierung aufeinanderliegend, ein gutes Modell für eine Membran abgeben würden.
Die hydrophoben Anteile wären demnach nach innen gerichtet, die hydrophilen nach außen. 1935 griffen H. DAVSON und J. F. DANIELLI die Idee auf und postulierten, daß die Proteine dieser Doppellipidschicht (einem bimolekularen Film) aufgelagert seien und dabei mit den hydrophilen Köpfen der Phospholipide in Wechselwirkung treten sollten.

Membranmodell nach DAVSON und DANIELLI. Das Modell sieht vor, daß die Lipide eine Doppelschicht ausbilden und daß die Proteine dem Lipidfilm aufgelagert sind. Die beiden Modelle (linke und rechte Darstellung) sind Varianten, in denen unterschiedliche Konformationen der Proteine angenommen werden.
Das Modell ging als DAVSON-DANIELLI-Modell in die Literatur ein und hatte 37 Jahre Bestand. Es erklärte zwar ganz richtig die Organisation der Lipidmoleküle, doch konnte es die zahlreichen Eigenschaften der Membranproteine nicht widerspruchsfrei deuten.
1972 wurde es durch das Fluid mosaic model von J. S. SINGER und G. NICOLSON (University of California, San Diego) abgelöst.
Dieses Modell beschreibt die Membran als ein flüssiges Mosaik;
es sieht vor, daß die Lipide ein visköses zweidimensionales
Lösungsmittel bilden, in das Proteine mehr oder weniger tief
eingelassen und in dem sie verankert (integriert) sind.
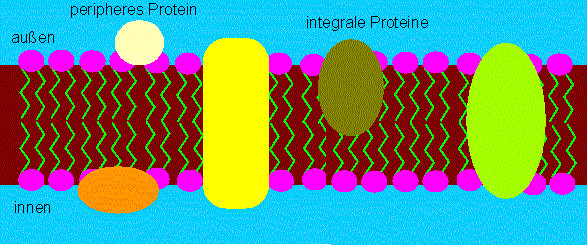
Fluid mosaic model der Membranstruktur. Die Proteinmoleküle sitzen im, nicht auf dem Lipidfilm. Manche ragen durch die Lipidschicht hindurch. Die hydrophoben Bereiche stehen in Kontakt mit den hydrophoben Schwänzen der Lipidmoleküle. Die polaren Gruppen ragen nach außen.(Nach S. J. SINGER und G. L. NICOLSON, 1972).
Proteine dieser Kategorie bezeichnet man als integrale Membranproteine. Manche von ihnen reichen von der einen zur anderen Membranseite, viele sind zu Aggregaten vereint, und es ist daher leicht vorstellbar, wie sich Kanäle oder Poren durch die Membran hindurch bilden können. Das Modell macht aber auch die Vorhersage, daß sich die Moleküle in der Membranebene (lateral) frei bewegen können. Dem scheint zwar die Beobachtung zu widersprechen, daß bestimmte Proteine in der Membran oft nicht statistisch verteilt sind und daß eine Membran wie ein Mosaik aus Bereichen mit unterschiedlicher Funktion zusammengesetzt ist. Dennoch stehen auch diese Erscheinungen mit dem Modell im Einklang, wenn man davon ausgeht, daß Protein-Proteininteraktionen vorhanden sind, die die freie Beweglichkeit der einzelnen Moleküle beeinträchtigen. Zudem ist bekannt, daß es außer den integralen Membranproteinen assoziierte (periphere) Proteine gibt, die der Membran aufgelagert oder ihr unterlagert sein können, die auch selbst untereinander in Wechselwirkung stehen. Oft sind Proteinanteile, die mit der wässrigen Phase in Kontakt stehen, glykosyliert.
| Porin aus Rhodobacter capsulatus | Porin aus Rhodpseudomonas blastica | PhoE aus Escherichia coli |
Der Stoffaustausch durch die Membranen wird durch hochspezifische Transportsysteme bewerkstelligt. Hierzu geh�ren die oben abgebildeten, bei Bakterien untersuchten Porine. In der Bildserie sind Porine von verschiedenen Arten dargestellt. Wie ersichtlich, �hneln sich die einzelnen Strukturen, was auf Homologie hinweist und als Beleg daf�r zu gelten hat, da� es sich um sehr konservative Proteine handelt, also solche, die sich im Verlauf der Evolution nur wenig ver�ndert haben. Die Abbildungen m�gen zu dem Trugschlu� verleiten, Poren in Membranen seien sehr locker strukturierte Gebilde. Benutzen Sie die rechte Maustaste und w�hlen Sie die Optionen: Display / Spacefill / Van der Waal. Weitere an Poren und Kan�len beteiligte Proteine sind nachfolgend dargestellt:
Die spezifische Form bestimmter Zellen, z.B. der Erythrozyten (roten Blutkörperchen), wird durch ein solches Proteinskelett determiniert, über das die Membran gespannt ist.
Die einzelnen Membrantypen einer Zelle unterscheiden sich aufgrund unterschiedlicher Zusammensetzung:
Unterschiedliche Phospholipide (und andere Lipide, z.B. Cholesterol) in den beiden Schichten des Lipidfilms. Hierdurch wird eine Asymmetrie hervorgerufen. Die Unterschiede können qualitativer und auch quantitativer Natur sein.
Unterschiedliche Lipidzusammensetzung in Membranen unterschiedlicher Herkunft.
Unterschiedliche Proteine bedingen unterschiedliche Funktionen von Membranen. Manche Enzymaktivitäten können als Marker für bestimmte Membrantypen herangezogen werden.
Wichtig ist auch die Orientierung bestimmter Proteine in der Membran.
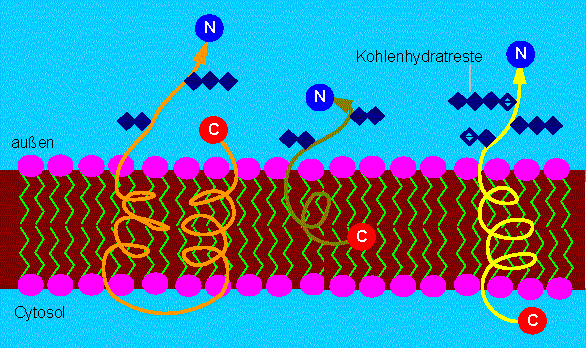
Unterschiedliche Möglichkeiten der Verankerung integrierter Membranproteine in der Membran. Das N-terminale Ende befindet sich stets an der dem Cytosol abgewandten Seite. Vielfach tragen Proteine am N-terminalen Ende Kohlenhydratreste. Typische Membranproteine können daher der Klasse der Glykoproteine zugeordnet werden (es gibt jedoch auch Ausnahmen). Das C-terminale Ende kann, wie die Abbildung zeigt, ebenfalls außen, in der Lipidschicht oder an der Cytosolseite (dem Syntheseort des Proteins) lokalisiert sein.
Membranen üben zahlreiche Funktionen aus. Die eingangs aufgezählten Kriterien können weiter aufgeschlüsselt werden:
Permeabilität (Durchlässigkeit) für einen Molekültyp und Impermeabilität (Undurchlässigkeit) für einen anderen gehörten zu den ersten Eigenschaften, die auf das Vorhandensein von Membranen schließen ließen. Alle biologischen Membranen sind selektiv permeabel.
Membranen trennen Reaktionsräume, die sich aufgrund ihrer chemischen Zusammensetzung, des pH-Werts und des elektrischen Potentials voneinander unterscheiden können. Es kann daher sowohl ein pH- als auch ein elektrischer Gradient durch eine Membran hindurch gemessen werden.
Manche Ionen oder kleine Moleküle (Substrate) können unter Energieverbrauch durch Membranen hindurch befördert werden. Man spricht hierbei von einem aktiven Transport, im Gegensatz zu einem passiven, der in Richtung eines Konzentrationsgradienten erfolgt.
Aufgrund unterschiedlicher Zusammensetzung nehmen die einzelnen Membrantypen grundsätzlich voneinander verschiedene Funktionen wahr. Hochgradig spezialisierte Membranen, wie die Thylakoidmembranen der Chloroplasten und die innere Membran der Mitochondrien, sind an Energieumwandlungen beteiligt.
Um passiven Transport zu verstehen, müssen wir uns mit den Themen Diffusion, Permeabilität und Osmose auseinandersetzen. Zum Verständnis des aktiven Transports gehört die Kenntnis der Struktur und Funktion integraler Proteine und der elektrochemischen Eigenschaften der Membran und zum Verständnis von Energieumwandlungen (Energietransformationen) die Kenntnis aller in der betreffenden Membran enthaltenen Moleküle.
Unabhängig hiervon stehen die Betrachtung der einzelnen Kompartimente und die Fragen: Welche Reaktionen finden in ihnen statt, welche Bedeutung kommt der räumlichen Trennung einzelner Biosyntheseketten zu, und wie sieht der Stoff- und Energieaustausch zwischen den einzelnen Kompartimenten aus? Wie wird Information ausgetauscht? Wie werden unterschiedliche Ionen- und Metabolitkonzentrationen aufrechterhalten? Wie gelangen große Moleküle (z.B. Proteine) durch Membranen hindurch, und schließlich, wie werden Membranen gebildet; können sie ineinander übergehen, wie würde dann ein Membranfluß aussehen und welchen Einfluß übt er auf den Molekültransport innerhalb der Zellen, von Zelle zu Zelle und zwischen Zellen und ihrer Umwelt aus?
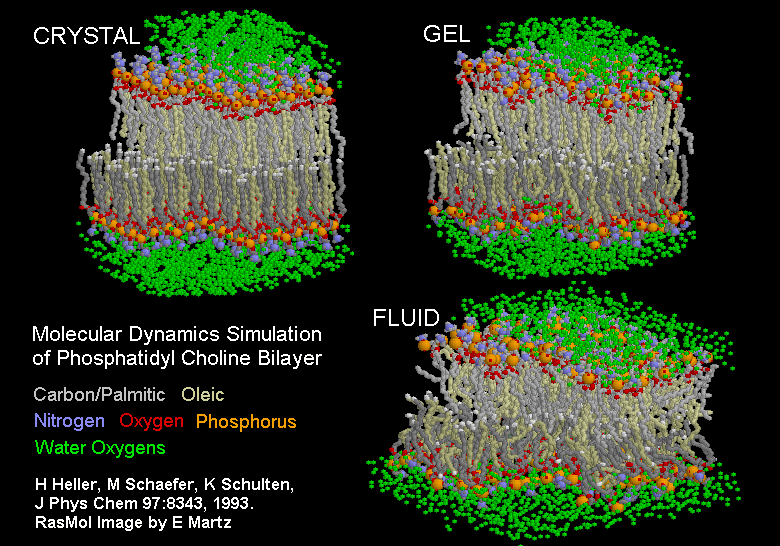
|
|