
Atome, Ionen und Moleküle sind in ständiger Bewegung. Sie stoßen dabei untereinander zusammen und werden dadurch aus ihrer Bahn geworfen. In einem idealen Gas führen die einzelnen Partikel ungehinderte, d.h. voneinander unabhängige Bewegungen aus. Die Partikelverteilung bleibt dabei unbeeinflußt, es gibt daher keine vorherrschende oder bevorzugte Bewegungsrichtung. Die Geschwindigkeit ist temperaturabhängig, mit steigender Temperatur zunehmed. Man spricht daher von thermischer Bewegung (T). In Flüssigkeiten sind die Bewegungen eingeschränkt, doch unterliegt die Bewegung in einer echten Lösung den gleichen Kriterien wie in einem idealen Gas. Für viele Zwecke kann auch das Plasma einer Zelle wie eine solche Lösung beschrieben werden. Die Beschreibung läßt sich jedoch nicht auf alle Situationen ausdehnen (siehe Problem Kompartimentierung).
Wird eine konzentrierte Lösung einer beliebigen Substanz (z.B. eines Zuckers) mit Wasser überschichtet, bildet sich zunächst eine klare Trennfläche zwischen den beiden Flüssigkeiten aus. Da wir davon ausgegangen sind, daß gelöste Partikel (hier Zuckermoleküle) sich rein statistisch bewegen, werden zunächst einzelne, als f (t) ständig mehr die Trennfläche durchdringen. Einige von ihnen, doch nicht alle, werden wieder zurückkehren, so daß eine Nettobewegung (ein Nettofluß) von Teilchen in Richtung von Regionen, die ursprünglich frei von ihnen waren, entsteht.
Eine Bewegung von hoher zu niedriger Konzentration bezeichnet man als Diffusion. Erst nach gleichmäßiger Verteilung aller Partikel in einem System ist keine Nettobewegung mehr nachweisbar; der Konzentrationsausgleich ist erreicht, das System hat ein Gleichgewicht erreicht.
Wie läßt sich dieser Vorgang quantitativ beschreiben? Die Menge der Partikel, die eine bestimmte Entfernung durch Diffusion zurücklegen, hängt von der Querschnittsfläche des zu betrachtenden Gefäßes (F) und einer Stoffkonstanten (D) ab.

Damit läßt sich nun ein Nettofluß als Funktion der Zeit wie folgt beschreiben:
f(t) = DF (c1 - c2 / x1 - x0)
wobei c1 und c2 die Konzentrationen in den betrachteten Querschnittsflächen, und x1 - x0 der Abstand zwischen ihnen ist. Da die Konzentration mit wachsender Entfernung abnimmt, hat der Konzentrationsgradient einen negativen Wert. Schreibt man die obige Formel für beliebig kleine Zeiträume, müssen die Werte als Differentiale angegeben werden, und man kommt zu
dm / dt = DF (dc / dx ) [cm2 sec-1]
(= Ficksches Diffusionsgesetz).
Die Dimensionen der eingesetzten Größen sind: m: [Mol], F: [Quadratzentimeter], c: [Mol/Kubikzentimeter], x: [cm].
Die Anzahl von Molen, die eine bestimmte Fläche pro Sekunde durchwandert, wird als Nettoflux (Phi) bezeichnet. Formelmäßig geschrieben:
Phi = - D (dc / dx) [Mol cm-2 sec-1]
oder
Phi = - D (cl - c2 / x1 - x0)
nach D aufgelöst:
D = -Phi / (cl - c2 / x1 - x0)
Die Größe (c1-c2) / (x1-x0) wird als Konzentrationsgradient (oder Konzentrationsgefälle) bezeichnet, D ist die Diffusionskonstante, die als die Menge einer Substanz (in Mol), welche pro Zeiteinheit (sec) durch eine Flächeneinheit (Quadratzentimeter) bei einem Konzentrationsgradienten von 1 (Mol/cm) diffundiert, definiert ist.
Die Diffusion erfolgt über kurze Entfernungen sehr schnell, ist aber für große Entfernungen extrem langsam. Die Entfernung geht ja im Quadrat in die Gleichung ein und ist proportional der zur Verfügung stehenden Fläche. Diffusion ist wichtig, wenn wir Molekülbewegungen innerhalb von Zellen oder zwischen benachbarten Zellen betrachten. Sie verliert jedoch an Bedeutung, wenn wir ganze Gewebe untersuchen, und ist völlig bedeutungslos, wenn wir z.B. an den Transport von Assimilaten aus Blättern in die Wurzeln denken.
Unter Permeabilität versteht man die Diffusion von Partikeln durch Membranen (Grenzflächen) hindurch. Dabei ist es zunächst belanglos, ob wir natürliche oder künstliche Membranen (z.B. Plastikfolien) betrachten. Um eine quantitative Aussage zu erhalten, nehmen wir in grober Näherung an, die Membran sei eine Lösungsmittelschicht der Dicke d.
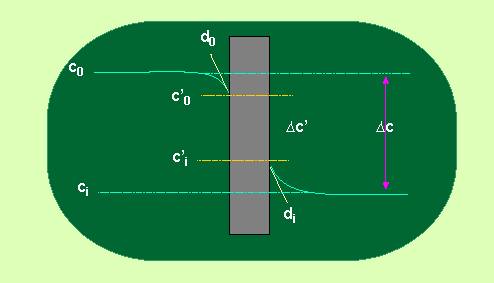
Unter dieser Annahme ist der Durchfluß durch eine Membran der Diffusionskonstanten direkt proportional. Sie ist durchweg niedriger als im Wasser. Der Nettoflux durch die Membran kann demnach als
Phi = - D (c1 - c2 / d) = - D / d (c1 - c2)
beschrieben werden, wobei der Ausdruck D/d die Permeabilitätskonstante ist und die Dimension [cm/sec] hat.
Biologische Membranen sind nicht für alle Substanzen gleich gut durchlässig. Sie sind selektiv permeabel, d.h. die Membranen sind für eine Substanz A permeabel, für eine Substanz B impermeabel. Strenggenommen sind sie nur für Wasser und einige Gase wie Sauerstoff, Stickstoff, Kohlendioxyd u.a. durchlässig, denn nur diese können eine Membranbarriere durch freie Diffusion überwinden. Wie oben abgeleitet, folgt diese ausschließlich den Gesetzen der Thermodynamik. Moleküle zeichnen sich durch spezifische Eigenschaften wie Molekülgröße, Ionisierbarkeit oder Löslichkeit in einer Membran aus. Solche lipophilen (fettlöslichen) Moleküle können sie daher eher passieren als die hydrophilen.
Membranen enthalten in ihrer Lipiddoppelschicht noch eine Reihe weiterer integraler Komponenten, von denen für uns hier die Proteine am wichtigsten sind. Durch eine spezifische Anordnung von Membrankomponenten können Poren oder Kanäle entstehen, durch die bestimmte Ionen und Moleküle selektiv durch die Membran diffundieren können. Ihre Durchlässigkeit hängt damit zum einen vom Porendurchmesser ab, zum anderen aber auch von seinem Ladungszustand. Kleine Anionen wie das Chlorid können durch positiv geladene Poren leicht passieren. Kationen werden zurückgehalten. Bei negativ geladenen Poren liegen die Verhältnisse umgekehrt. Man spricht hier von eingeschränkter Diffusion. Etliche kleine Moleküle, wie Zucker und Aminosäuren werden unter bestimmten Voraussetzungen scheinbar bevorzugt durch eine Membran hindurchgeschleust. Die Permeationskinetik folgt aber nicht der obigen Formel, vielmehr ähnelt sie einer Enzym-Substrat-Umsatzkinetik. Hier haben wir es mit einer erleichterten (oder geförderten) Diffusion zu tun. Die Durchlässigkeit ist selektiv und von der Anwesenheit spezifischer membrangebundener Träger- oder Carriermoleküle mit einer Affinität zu einer begrenzten Gruppe chemisch verwandter Substanzen abhängig. Es sind demnach nicht die Substanzen (Substrate) selbst, die die Membran passieren, sondern ein Substrat-Carrier-Komplex. Zusammenfassend kann erleichterte Diffusion durch folgende Kriterien beschrieben werden:
- Die Membran enthält spezifische Träger- oder Carriermoleküle. Man spricht daher auch von trägervermitteltem Transport.
Die Carrier (meist Proteine, aber auch bestimmte Antibiotika wie das Valinomycin) binden das zu transportierende Substrat, z.B. einen Zucker oder eine Aminosäure. Im Gegensatz zu Enzymen wird es nicht umgesetzt, sondern durch die Membran translociert.
Mehrere Substrate können um den gleichen Carrier konkurrieren. Ihre Bindungseigenschaften zu ihm können unterschiedlich sein; folglich sind auch die Transportkinetiken substratspezifisch.
Der beladene Carrier kreuzt die Membran mit einer anderen Geschwindigkeit als der unbeladene.allein der Konzentrationsgradient der zu transportierenden Substanz.
Die treibende Kraft der erleichterten Diffusion ist wie bei einfacher Diffusion
Die maximale Transportgeschwindigkeit hängt von der Anzahl der in der Membran vorhandenen Carriermoleküle ab.
|
|