

Die Gymnospermen wurden erstmals von THEOPHRAST erwähnt. Er faßte darunter alle Pflanzen zusammen, deren Samen ungeschützt sind. Es sind primitive Samenpflanzen, jedoch repräsentieren die Gymnospermae kein phylogenetisch einheitliches Taxon. Die rezenten Arten können zwei großen Unterabteilungen, den Coniferophytinae und den Cycadophytinae zugeordnet werden. Es ist viel darüber debattiert worden, ob sie diphyletischen Ursprungs sind oder doch auf gemeinsame Vorfahren zurückgeführt werden können.
Heute stellt sich die Sache so dar, daß beide als Nachfahren einer Pteridophytengruppe, den Progymnospermae anzusehen sind. Aber: die Progymnospermae sind in sich ein heterogenes (polyphyletisches) Taxon, und es sieht so aus, als würden sich die Coniferophytinae und Cycadophytinae von unterschiedlichen Formen ableiten.
Die Progymnospermae traten im Devon in Erscheinung. Sie sind echte Pteridophyten, denn sie vermehren sich durch Sporen. Homosporie schien vorherrschend zu sein, wenngleich bei einigen Formen Heterosporie gefunden wurde. Es gibt Hinweise darauf, daß sich von ihnen nicht nur die Spermatophyta, sondern auch die echten Farne (Pteropsida) ableiten. Sie selbst entstammen vermutlich den Trimerophytatae, von denen sie sich durch ein komplexeres Leitungssystem und sekundäres Dickenwachstum unterscheiden.
Die primitiveren Formen rechnet man dem Aneurophyton-Typ zu, der durch ein dreidimensionales Verzweigungsmuster der Sprosse und mit Markstrahlen durchsetztes, poröses Holz gekennzeichnet ist. Der abgeleitete Archaeopteris-Typ besaß meist gefiederte Wedel und poröses oder kompakt strukturiertes Holz; progressive Vertreter waren heterospor. Die Holzanatomie war der entscheidende Anlaß zu der Annahme, daß die Gymnospermen von den Progymnospermen abstammen, wobei die Coniferophytinae vermutlich aus Formen mit festem Holz und weniger deutlich ausgeprägten Blättern, und die Cycadophytinae aus wedeltragenden Formen mit durch Parenchym aufgelockertem Holz hervorgegangen sind.
Aus der eben vorgetragenen Argumentation wird deutlich, daß die Verholzung von Haupt- und Nebentrieben der Vervollkommnung der Fortpflanzungsorgane vorausging. Alle rezenten und fossilen Gymnospermen (und Angiospermen) sind blüten- und samenbildend. Unreifer Pollen ist den Mikrosporen, die frühen Stadien der Samenanlage den Megasporen der Pteridophyten homolog. In späten Entwicklungsstadien sind der nunmehr reife Pollen dem männlichen Gametophyten, der Embryosack (ein Teil der Samenanlage) dem weiblichen Gametophyten homolog. Die Homologie ließ sich an Hand zahlreicher fossiler Belege lückenlos dokumentieren. Zur Veranschaulichung sei zunächst die Entwicklung der Samenanlage und des Pollens an einem Modellbeispiel erläutert, aus dem die Homologisierung einzelner Teile und deren Benennungen hervorgehen:
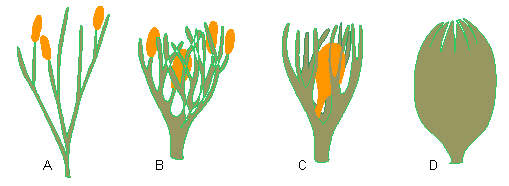
Sukzessiv aufeinanderfolgende Schritte, die zum Einschluß eines Sporangiums durch Nucellus und Integument führten, so wie es bei den Samen der Pteridospermen in Erscheinung tritt. A. terminal sitzende Sporangien (Rhynia), B. Übergipfelung eines Sporangiums durch fertile und sterile Telome (Hedeia corymbosa). C. ein hypothetisches Stadium. Ein vergrößertes Sporangium, das von einem Telomnetzwerk eingeschlossen wird. D. Samen mit Sporangium (Nucellus), von verwachsenen Telomen (= Integument) umgeben (Nach WALTON, 1940, aus H. N. ANDREWS, 1961).
Reife Megasporen (der Pteridophyta) werden von der Mutterpflanze abgestoßen und entwickeln sich an einem neuen, geeigneten Standort zu weiblichen Gametangien. Die frühen Stadien der Samenanlage (der Spermatophyta), den frühen Stadien des Mega-Gametophyten homolog, bleiben mit der Mutterpflanze verbunden. Der Nucellus und sein Inhalt entwickeln sich im Schutz eines speziellen Organs, dem Integument. Es entstand durch Verwachsung ursprünglich sporangientragender, dann steril gewordener Sproßabschnitte (Telome).
Reifer Pollen und Embryosack sind Gametophyten, sie beinhalten männliche und weibliche Gametangien (Antheridium und Archegonien), bzw. das was von ihnen noch an Zellen oder Kernen übrig ist
Reife Megasporen werden (bei Pteridophyten) von der Mutterpflanze abgestoßen und entwickeln sich an einem neuen geeigneten Standort zu weiblichen Gametophyten. Bei den Gymnospermen entwickelt sich die Embryosackmutterzelle zum Embryosack, der den weiblichen Gametophyten darstellt. Hier findet die Meiose statt, und in der Regel entwickelt sich nur eine der vier haploiden Tochterzellen weiter. Die drei übrigen gehen bei den Gymnospermen im Verlauf der Zeit zugrunde. Der Zellkern der zur weiteren Entwicklung determinierten Zelle durchläuft eine Vielzahl von Kernteilungen. Es entsteht dadurch ein Synzytium (ein vielkerniges Gebilde). Im Anschluß an die Kernteilungen werden Zellwände eingezogen. Die Zellwandbildung geht von der Peripherie des Synzytiums aus.
Im sich entwickelnden Embryosack differenzieren sich einige der am apikalen Pol gelegenen Zellen zu Archegonien, die aus einer zentral gelegenen Eizelle, einer Bauchkanalzelle und Halszellen, die den Halswandzellen des Pteridophyten-Archegoniums homolog sind. Ein Kranz kleiner Zellen wird vom Prothallium beigesteuert, ist aber nicht den Halswandzellen der Pteridophyten-Archegonien homolog
Während sich die Archegonien bilden, setzt das Integument sein Wachstum so lange fort, bis es die Samenanlage fast vollständig umschlossen hat. Die apikal verbleibende kanalförmige Öffnung wird Mikropyle genannt. Zwischen dem Integument und dem Nucellus bildet sich eine mehr oder weniger große Pollenkammer, deren Ausdehnung artspezifisch ist; ihre Bildung ist, z.T. wenigstens, auf einen Zerfall apikalliegender Zellen des Nucellus zurückzuführen. Diese Exposition ist ein charakteristisches Merkmal der Gymnospermen.
Wie erwähnt, entspricht ein unreifes Pollenkorn einer Mikrospore. Pollen wird in Pollensäcken erzeugt, welche ihrerseits den Mikrosporangien der Pteridophyten entsprechen. Reife Pollenkörner sind mehrzellig. Sie sind stets von einer dicken mehrschichtigen Hülle umgeben, deren Struktur und Zusammensetzung als artdiagnostisches Merkmal verwendbar ist. Durch Wind verbreitete Pollenkörner besitzen oft einen oder mehrere Luftsäcke. Aus den zentral gelegenen Zellen entwickelt sich ein Antheridium und darin zwei männliche Gameten. Nur bei den Cycadophytinae (Ausnahmen: Gnetales) und Ginkgo kommen begeißelte Gameten vor, bei allen übrigen Gymnospermen und bei den Angiospermen sind sie unbegeißelt.
Die erste Voraussetzung für einen Befruchtungsvorgang ist das Eindringen von Pollen durch die Mikropyle in die Pollenkammer, die zweite, das Auswachsen eines Pollenschlauchs, in den (bei den Coniferophytina) der männliche Gametenkern einwandert und der mit der Eizelle Kontakt aufnimmt. Nach der Fusion der beiden Zellen kommen männlicher und weiblicher Kern zusammen und verschmelzen zu einer Zygote. Bei den Cycadophytinae dient der Pollenschlauch ausschließlich der Ernährung des männlichen Gametophyten.
Durch den Befruchtungsvorgang ist der erste Teil des Samenbildungsprozesses abgeschlossen. Es folgen die einleitenden Schritte einer Embryonalentwicklung, und schließlich die Freisetzung des reifen Samen.
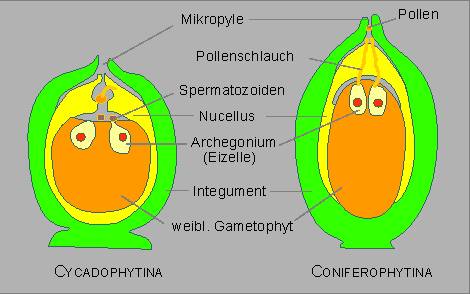
Samenanlagen bei Gynmospermen (Nach W. ZIMMERMANN, 1959)
Die Coniferophytinae und die Cycadophytinae unterscheiden sich in der Struktur des Holzes, der Blattform 5nd der Samensymmetrie. Das Holz der Coniferophytinae ist fest, denn es ist von nur wenigen dünnen, parenchymatischen Markstrahlen durchsetzt; hingegen ist das der Cycadophytinae sehr porös, weil es durch sehr breite parenchymatische Markstrahlen aufgelockert ist. Coniferophytinenholz wird wirtschaftlich genutzt (Kiefer, Fichte usw.), Cycadophytinenholz hat keinen wirtschaftlichen Nutzen.
Die Blätter (Nadeln) der Coniferophytinae können, von einer Ausnahme abgesehen (Ginkgoales), als Mikrophylle klassifiziert werden, denn sie werden durch nur eine oder zwei parallel liegende Blattnerven versorgt. Cycadophytinenblätter sind, wie Farnwedel, Megaphylle. Die Blattspreiten enthalten ein umfangreiches, durch Anastomosen untereinander verknüpftes Adersystem. Coniferophytinensamen sind bilateral gebaut, Cycadophytinensamen in der Regel radiärsymmetrisch.
Die Untergliederung in Ordnungen sieht wie folgt aus:
Die Organisation der Vegetationskörper großer, komplex strukturierter Pflanzen ist aus fossilen Fragmenten oft nur sehr schwer zu rekonstruieren, da die meisten entweder nur Teile des Stamms, Teile der Blätter, der Reproduktionsorgane oder Samen enthalten. Anhand eines Blattfragments ohne Samenanlage oder ohne Sporangium kann man daher nicht einmal entscheiden, ob es von einem Farn oder von einem Samenfarn stammt. Viele der Schlußfolgerungen und Rekonstruktionsversuche beruhen daher auf statistischen Auswertungen, bei denen die Häufigkeit, mit der bestimmte Teile verschiedener Organe in der gleichen Formation auftreten, als Entscheidungshilfe dient.
© Peter v. Sengbusch - b-online@botanik.uni-hamburg.de