

"Pink Lagune" von Qalah (Oman).Die Färbung des Wassers (Salzwasser) der Lagune (an der Arabischen See) beruht auf dem massenhaften Vorkommen einer volvocalen zweigeißligen Grünalge. Die Rotfärbung beruht auf der Bildung eines Carotinoids. In Kultur verlieren die Algen diese Fähigkeit und sehen dann ganz normal grün aus.
Die Grünalgen scheinen den übrigen grünen Pflanzen näher zu stehen als den anderen Abteilungen der Algen. Beide Gruppen besitzen die gleichen Photosynthesepigmente (Chlorophyll a und b), den gleichen Satz an Carotinoiden (alpha-, beta- und gamma-Carotin, Lutein, Zeaxanthin, Violaxanthin u.a.), den gleichen Reservestoff (Stärke) und die gleiche Gerüstsubstanz der Zellwand: Cellulose. Es lag daher nahe, die Chlorophyta mit allen zur Verfügung stehenden analytischen Methoden zu untersuchen, um Näheres über ihren phylogenetischen Hintergrund zu erfahren; insbesondere über ihre Beziehung zu den Landpflanzen. Viele der bisherigen Bemühungen schlugen fehl, doch in den letzten Jahren bahnt sich aufgrund ultrastruktureller Untersuchungen ein Fortschritt auch auf diesem Gebiet an. So wurde dem Bau der begeißelten Zellen zunehmend Bedeutung beigemessen. Als besonders aufschlußreich erwies sich die Analyse des Cytoskeletts und des Bewegungssystems. Man fand, daß die Komponenten des Geißelapparates in Struktur und Anordnung sehr verschieden und für bestimmte phylogenetische Linien charakteristisch sind.
Das gleiche gilt auch für die mit Mitose und Cytokinese verbundenen Strukturen. Während der Mitose (höherer Organismen) löst sich die Kernmembran auf. Doch bei einer Anzahl von Grünalgen (und vielen Protisten) bleibt die Kernhülle auch während der Zellteilung in unterschiedlichem Grade erhalten, chromosomenähnliche Aggregate treten innerhalb des membranumschlossenen Volumens in Erscheinung. Die Mikrotubuli der Telophase können unterschiedlich lange persistieren.
Schließlich gibt es unterschiedlichste Formen der Zelldurchschnürung und Querwandbildung. Während sich bei der Zellteilung der Landpflanzen ein Phragmoplast ausbildet, bei dem sich die Mikrotubuli senkrecht zur Teilungsebene organisieren, fand man zunächst bei etlichen Grünalgen einen "Phycoplasten", der aus parallel zur Äquatorialebene gelagerten Mikrotubuli besteht. Inzwischen kennt man eine ganze Reihe (ca. 8) verschiedene Mitose- und Zellteilungstypen. Auch der Phragmoplast ist darunter und gibt natürlich Anlaß zu Spekulationen über den Ursprung der Landpflanzen.

Phycoplast und Phragmoplast bei verschiedenen Grünalgen. A: Fritschiella; B: Chlamydomonas; C: Coleochaete; D: Klebsormidium. Beim Phycoplasten sind die Mikrotubuli parallel, beim Phragmoplasten senkrecht zur Teilungsebene angeordnet (Nach P. H. RAVEN, R. F. EVERT, H. CURTIS, 1981; und G. L. FLOYD).
Aufgrund von DNS-Sequenzierung ausgewählter Gene (besonders solche der ribosomalen Untereinheiten) zeichnen sich in letzter Zeit neue phylogenetische Zusammenhänge ab, die zu einer erheblichen Umstellung des herkömmlichen Systems der Grünalgen führen. Da aber noch zahlreiche Widersprüche zu klären sind, ist es zu früh, heute ein neues, verbindliches System vorzustellen. Aus pragmatischen Gründen können die Chlorophyta unter Berücksichtigung der genannten und anderer ultrastruktureller Kriterien fünf Klassen zugeordnet werden:
Die genannten Klassen sind in sich zum Teil sehr heterogene Gruppierungen. Es gibt daher noch zahlreiche Unstimmigkeiten bezüglich der Zuordnung einzelner Gattungen zu höheren Taxa oder der Beziehungen dieser Taxa (Ordnungen, Klassen) zueinander. Man hat früher immer wieder versucht, aufgrund von morphologischen Ähnlichkeiten Verwandtschaften abzuleiten. Heute kann man z.B. innerhalb der von PASCHER definierten "Organisationsstufen" nur noch konvergente Gruppen beschreiben. Die "Stufen" wiederholen sich parallel in verschiedenen Taxa. Vermutlich gilt hier das gleiche, was für die Angiospermen gesagt wurde: Das Genom enthält mehr genetische Information, als für die Existenz der Individuen (Arten) benötigt wird. Durch Umstrukturierung können daher in den verschiedensten phylogenetisch nicht zusammenhängenden Gruppen gleichartige Merkmale in Erscheinung treten. Für die Wahrscheinlichkeit dieser Annahme gibt es verschiedene Hinweise.
Die Mehrzahl der Chlorophyta ist stets oder zeitweise begeißelt (zu den Ausnahmen gehören u.a. die Zygnematales). Es sind zwei oder mehr gleich lange (isokonte) Geißeln vorhanden. Begeißelte Stadien (vegetative Zellen, Zoosporen, Gameten) bewegen sich phototaktisch, viele Arten besitzen ein Stigma ("Augenfleck"), das in der Regel in einem Chloroplasten lokalisiert ist. Augenflecken kommen auch bei nicht-begeißelten Arten, so z.B. bei Spirotaenia condensata vor. Wie schon an anderer Stelle dargelegt, besitzen die Grünalgen meist viele und unterschiedlich strukturierte Chloroplasten
Grünalgen sind weit verbreitet. Die meisten Arten findet man im Süßwasser, andere im Salz- und/oder Brackwasser, im Boden, an Baumstämmen oder als Symbionten von Protozoen (Paramaecium bursaria), Coelenteraten (Hydra) und Pilzen (Flechten: Lichenes). Die Mehrzahl der nichtaquatischen Arten ist einzellig. Nur wenige - volvocale - Arten (s. Titelbild) erreichen lokal so hohe Individuenzahlen wie die Cyanophyta, Euglenophyta oder Bacillariophyceae.
Sie sind daher auch nur selten an Wasserblüten beteiligt. Einige fädige Arten (z. B. Spirogyra-, Mougeotia-, Enteromorpha- und Cladophora-Arten) bilden, wo sie in Massen auftreten, sogenannte Watten aus, die aus einer Vielzahl ineinander verflochtener Fäden bestehen. Solche, meist an Oberflächen schwimmende Aggregate sind ein Kennzeichen stark eutropher Gewässer.
In einigen Gruppen der Grünalgen kommen spezifische, bizarr aussehende Zellformen vor. Sie treten vor allem bei Einzellern auf und sind bei den Desmidiaceen, einer Zygnematales-Familie, besonders auffällig. Viele planktisch lebende Arten (Chlorococcales) sind mit Schwebefortsätzen versehen. Die meisten Arten sind von mehr oder weniger voluminösen Gallerten unterschiedlicher chemischer Zusammensetzung und Konsistenz umgeben, die dem Zusammenhalt der Individuen, der Anheftung an die Unterlage, dem Stoffaustausch und der Bewegung dienen können. Für viele Bakterien bilden sie einen bevorzugten Lebensraum. Zellen der Chlorophyta sind in der Regel einkernig und haploid, selten sind sie mehrkernig (polyenergid). Die bei sexueller Fortpflanzung in Erscheinung tretenden Gameten sind oft isogam; Anisogamie als abgeleitete Erscheinung kommt vor, ebenso Oogamie (letztere bei primitiven und bei spezialisierten Gattungen: Chlamydomonas, Oedogonium. Chara, u.a.).
Es besteht keine einhellige Meinung über die Anzahl der Ordnungen. In der folgenden Zusammenstellung sind die wichtigsten, und damit die am eindeutigsten eingegrenzten, wiedergegeben:
Volvocales: Die Volvocales enthalten einzellige und koloniebildende Arten. In der Regel sind die Zellen begeißelt. Einige Arten können jedoch unter bestimmten Voraussetzungen zeitweilig in ein unbegeißeltes "Palmella"-Stadium übergehen. Die Zellen der Volvocales sind meist von einer mehrschichtigen, protein- und kohlenhydrathaltigen Wand umgeben.
Die Lage und Form des Chloroplasten ist artspezifisch. Typisch ist der
becherförmige Chloroplast von Chlamydomonas, der ein großes
Pyrenoid enthält und den Zellkern umgibt. Im vorderen Bereich des
Chloroplasten, dicht unter Plastidenhülle und Plasmalemma liegt das
karotinoidhaltige Stigma. Der eigentliche Photorezeptor wird aber von den
peripher dazu liegenden Membranen gebildet.
Links: Durch Haematococcus pluvialis gefärbtes Wasser in einer Pfütze (Aufn.: K. HANDKE), rechts: Volvox aureus

Auffallend ist der zentral gelegene, mit Fortsätzen versehene Chloroplast von Stephanosphaera. Unter den Volvocales treten oft Formen mit erhöhtem Carotinoidgehalt auf: Chlamydomonas nivalis verursacht Rotfärbung von Schnee ("Blut"), Haematococcus pluvialis eine rote Verfärbung von Regenpfützen. In Kultur kann Rotfärbung verschiedener Arten durch Stickstoffmangel hervorgerufen werden (Bildung von Sekundärcarotinoiden).
Die vorn gelegenen isokonten Geißeln durchdringen eine typische Wandpapille. Sie schlagen synchron nach dem "Ruderschlagprinzip": ausgestreckt nach hinten, mit einem von der Basis zur Spitze laufenden Knick nach vorn, so wird verhindert, daß die Zellen beim Schlag nach vorn wieder zurücktreibt. Am apikalen Pol liegen ferner die pulsierenden Vakuolen, die sich alternierend füllen und entleeren.
Bei der vegetativen Fortpflanzung verläuft die Trennung der Tochterzellen vom vorderen zum hinteren Pol. Mehrere Teilungen können innerhalb der Mutterzellwand aufeinander folgen, wobei es zur Umorientierung der Achsen kommt. Nach der Begeißelung werden die Tochterzellen durch enzymatischen Abbau der Mutterzellwand freigesetzt.
Die stets einzelligen Arten aus der Gattung Chlamydomonas gehören zu den vielgenutzten Objekten pflanzlicher Grundlagenforschung. Besondere Beachtung fanden die Kompatibilitätstypen (= Paarungstypen) von ansonsten isogamen Arten (+ und - Stämme). Vermerkt sei, daß es in der Gattung Chlamydomonas neben Isogamie auch Heterogamie und Ooogamie gibt.
Zu den Molekülen, die an der Spezifität der Paarungsreaktion (Verkleben [=Agglutination] der Geißeln zweier paarungsbereiter Gameten) beteiligt sind, gehören Glykokonjugate mit alpha-glykosidisch gebundenen Mannoseresten (L. WIESE und W. WIESE, 1975).
Arten der Gattung Carteria besitzen im Gegensatz zu denen von Chlamydomonas vier anstelle von zwei Geißeln.
Koloniebildende Gattungen: Die Kolonien von Volvocales sind meist Coenobien, d.h., sie enthalten (artspezifisch) stets eine charakteristische Zahl von Zellen. Diese wird bei der Ontogenese determiniert und kann nach der Ausdifferenzierung nicht mehr anwachsen. Alle Zellen sind stets die direkten Nachkommen einer Mutterzelle. In diese Gruppe gehören viele klassische Objekte des Botanikstudiums: Gonium, Pandorina, Eudorina, Volvox, Stephanosphaera u.a.
Die Gattung Gonium ist am einfachsten gebaut. Gonium sacculiferum besteht aus vier chlamydomonas-ähnlichen Zellen, die von einer gemeinsamen Gallerte umgeben sind. Die Coenobien von Gonium pectorale enthalten meist 16 Zellen, die anderer Arten 8, 16 oder 32.
Pandorina ist das bekannteste Beispiel für Coenobienbildung. Eine Kolonie enthält auch hier wieder meist 16 (-32) Zellen, und aus jeder geht wieder eine Kolonie mit 16 (- 32) Zellen hervor usw. Der Gattungsname entstammt der griechischen Mythologie. Die Götterbotin Pandora verbreitete Unheil, und jedem Unheil folgte neues Unheil.
Eine ähnliche Organisation finden wir bei Eudorina. Beide Gattungen unterscheiden sich aufgrund der Anordnung und der Zahl der Zellen im Coenobium. Pandorina-Zellen bilden einen kompakten Haufen,
Eudorina-Zellen sind in Form einer Hohlkugel angeordnet.
Volvox bildet riesige Kolonien, in denen 1000 bis 50 000 Zellen hohlkugelförmig vereint sein können. Der Koloniedurchmesser beträgt bis zu zwei Millimeter. A. van LEEUVENHOEK sah sie zum ersten Male im Januar 1700. Volvox-Kolonien weisen eine deutliche Differenzierung und Arbeitsteilung auf.
So wird der reproduktive Bereich zunehmend auf Zellen des hinteren Pols beschränkt. Die vorderen Zellen haben oft längere Geißeln und größere Stigmen. Alle Zellen reagieren phototaktisch, ihre Bewegungen sind durch den Lichteinfall synchronisiert. Es gibt daher eine bevorzugte Bewegungsrichtung der Kolonie. Zwischen den Zellen findet ein Informationsaustausch statt. Der sichtbare Ausdruck hiervon ist ein symmetrisch gebautes Netzwerk aus Plasmabrücken, durch die die Zellen untereinander in Verbindung stehen.
Man könnte daher durchaus von einem echten Vielzeller sprechen. Doch die Art, wie er entstanden ist, ist kein Modell zum Verständnis der Evolution der übrigen Vielzeller. Deren Entstehung verlief anders: einfacher und effizienter.
Zur Fortpflanzung gelangen reproduktive Zellen ins Kolonieinnere, wo aus ihnen durch zahlreiche aufeinanderfolgende Teilungen nach einem charakteristischen Schema Tochterkolonien entstehen. Die Geißeln der Tochterkoloniezellen sind anfangs ins Kolonieinnere gerichtet. Durch einen "Inversions"-Prozeß erlangen sie die richtige Orientierung. Die Freisetzung der Tochterkolonien vollzieht sich nach dem Reißen der schützenden Hülle der Mutterkolonie. Der Modus der sexuellen Fortpflanzung ist die Oogamie.
EINIGE BEMERKUNGEN ÜBER TETRASPORALES: Manche Autoren erheben dieses Taxon zu einer eigenständigen Ordnung, andere schlagen sie den Volvocales zu. Ein Kennzeichen vieler Arten ist die Anordnung der Zellen zu Vierergruppen innerhalb einer gemeinsamen Hülle. Tetrasporales repräsentieren sicherlich keine natürliche Gruppe, denn sie sind vornehmlich ein Sammelbecken von Arten auf einer bestimmten Organisationshöhe.
 Chlorococcales:
Die Chlorococcales sind dadurch charakterisiert, daß sie üblicherweise
unbegeißelt sind und feste Zellwände besitzen. In diesem Zustand
fehlen dann auch pulsierende Vakuolen und Stigmen. Gameten und Zoosporen
haben allerdings Geißeln und gleichen den reproduktiven Stadien der
Volvocales. Manche Arten (so z. B. Eremosphaera) bilden keine Zoosporen,
sondern innerhalb der Mutterzellwand Tochterzellen, die der Mutterzelle
gleichen (Autosporen).
Chlorococcales:
Die Chlorococcales sind dadurch charakterisiert, daß sie üblicherweise
unbegeißelt sind und feste Zellwände besitzen. In diesem Zustand
fehlen dann auch pulsierende Vakuolen und Stigmen. Gameten und Zoosporen
haben allerdings Geißeln und gleichen den reproduktiven Stadien der
Volvocales. Manche Arten (so z. B. Eremosphaera) bilden keine Zoosporen,
sondern innerhalb der Mutterzellwand Tochterzellen, die der Mutterzelle
gleichen (Autosporen).
Es gibt einzellige und koloniebildende, meist planktisch lebende Arten. Vielzellige Arten bestehen oft aus einer festgelegten Zahl von Zellen in typischer Anordnung (Scenedesmus, Pediastrum).
Diese Zellverbände entwickeln sich aus Tochterzellen, die sich voneinander lösen (als "Hemizoosporen", die begeißelt sind, oder als unbegeißelte "Sporen", die die Mutterzellwand zunächst nicht verlassen) und sich dann in typischer Weise arrangieren (Aggregationsverband). Diese Verbände wachsen heran, sprengen dann die Mutterzellwand und treten manchmal in einer spezifisch geformten Blase aus.
Die Zellwand der Chlorococcales besteht meist aus einem Cellulosegerüst, in das vielfach andere Moleküle eingelagert sind. Bei vielen Arten ist sie mehrschichtig, so besteht z.B. eine der Schichten der Chlorella- und Scenedesmus-Wand aus Polymeren aus ungesättigten Kohlenwasserstoffketten, die sich aus der Fettsäurebiosynthese herleiten lassen. Oft sind den Wänden strukturbildende Substanzen aufgelagert. Sexuelle Fortpflanzung ist selten und wenn, dann sind die Gameten isogam. In vielen Gattungen, z.B. Chlorella, wurde nie eine Gametenbildung gesehen.
Der bekannteste Vertreter der Chlorococcales ist Chlorella. Arten dieser Gattung sind Standardobjekte der Photosyntheseforschung. Sie sind im Süß- und Salzwasser sowie im Boden verbreitet. Die Zellen enthalten ein einziges, auffallend komplex strukturiertes Mitochondrium.
Zu den Verwandten von Chlorella gehört Prototheca mit ihren stets farblosen Plastiden. Chlorococcum ähnelt Chlorella morphologisch, bildet aber anstelle von Autosporen bewegliche Zoosporen aus. Die meisten Arten dieser Gattung vermehren sich ausschließlich asexuell, bei einer auf den Philippinen isolierten Art (Chlorococcum echinozygotum) wurde Isogamie nachgewiesen. Arten der Gattung Trebouxia leben oft in Symbiose mit Pilzen (Flechten).
Weitere weit verbreitete Gattungen sind Oocystis und Eremosphaera. Die Zellen vieler planktisch lebender Arten (Gattungen) sind mit Schwebefortsätzen versehen.
Koloniebildende Arten: Die meisten Chlorococcales-Kolonien können als Coenobien betrachtet werden. Typische Beispiele dafür finden wir in den Gattungen Scenedesmus, Ankistrodesmus und Pediastrum. Scenedesmus-Coenobien bestehen meist aus vier, seltener aus acht (oder 16) in Reihe nebeneinander angeordneter Zellen. Die außen liegenden tragen Schwebefortsätze. Isogamie wurde bei Scenedesmus obliquus nachgewiesen. Bei Ankistrodesmus sind die an den Enden spitz auslaufenden Zellen in Viererverbänden organisiert.
Pediastrum bildet zweidimensionale Coenobien, die je nach Art oder Kulturbedingung aus 8, 16, 32 oder 64 symmetrisch angeordneten Zellen bestehen. Während der Vermehrungsphase entsteht (wie bei Pandorina) aus jeder Zelle eine neue Kolonie. Bei sexueller Fortpflanzung werden wandlose Gameten gebildet, die, nach der Paarung, Zygotenbildung und Reduktionsteilung, durch mehrfach aufeinanderfolgende Teilungen zu einer neuen Kolonie heranwachsen. Fossil wurde Pediastrum im Perm und in der Trias nachgewiesen.
 Zu
den auffallendsten Chlorococcales gehört das Wassernetz Hydrodictyon.
Es besteht aus gestreckten Zellen, die an ihren Enden untereinander verbunden
sind und ein polygonales, in sich geschlossenes Netzwerk ausbilden, das
einen Durchmesser bis zu etwa einem Meter erreichen kann. Solche Wassernetze
in stehenden oder langsam fließenden Gewässern vorkommend, waren
schon den alten Chinesen bekannt, so daß man wohl sagen kann, Hydrodictyon
sei die erste beschriebene Alge. Die Zellen enthalten einen netzartigen
(retikulären), wandständigen Chloroplasten. Junge Zellen sind
einkernig, ältere meist vielkernig. Im Normalfall werden die in einer
Zelle gebildeten Zoosporen nicht freigesetzt, vielmehr verbinden sie sich
untereinander (unter Verlust ihrer Geißeln und Ausbildung einer Wand),
so daß bereits innerhalb einer Zelle ein neues Netz angelegt wird.
Während der sexuellen Fortpflanzung (Isogamie) wird ein polyedrisches,
zunächst einkerniges, später vielkerniges Zwischenstadium gebildet.
Zu
den auffallendsten Chlorococcales gehört das Wassernetz Hydrodictyon.
Es besteht aus gestreckten Zellen, die an ihren Enden untereinander verbunden
sind und ein polygonales, in sich geschlossenes Netzwerk ausbilden, das
einen Durchmesser bis zu etwa einem Meter erreichen kann. Solche Wassernetze
in stehenden oder langsam fließenden Gewässern vorkommend, waren
schon den alten Chinesen bekannt, so daß man wohl sagen kann, Hydrodictyon
sei die erste beschriebene Alge. Die Zellen enthalten einen netzartigen
(retikulären), wandständigen Chloroplasten. Junge Zellen sind
einkernig, ältere meist vielkernig. Im Normalfall werden die in einer
Zelle gebildeten Zoosporen nicht freigesetzt, vielmehr verbinden sie sich
untereinander (unter Verlust ihrer Geißeln und Ausbildung einer Wand),
so daß bereits innerhalb einer Zelle ein neues Netz angelegt wird.
Während der sexuellen Fortpflanzung (Isogamie) wird ein polyedrisches,
zunächst einkerniges, später vielkerniges Zwischenstadium gebildet.
Chaetophorales: Diese Ordnung muß aufgrund neuer Befunde wohl noch stärker eingegrenzt werden; sie überschnitt sich in älteren Systemen mit den Ulotrichales. In diese, möglicherweise nicht natürliche Ordnung werden Arten mit fädigem, z.T. verzweigtem Thallus gestellt. Einige Beispiele für Chaetophorales: Uronema, Chaetophora, Microthamnion, Aphanochaete, Draparnaldia, Fritschiella, Stigeoclonium.
Die Zoosporen tragen am Geißelpol eine Gallerte, die eine Polarität der Zelle markiert. Bei der Bildung von Zellaggregaten (nach Geißelabwurf) ist dieser Pol stets in Richtung des Zentrums orientiert. Die Verlängerung der Zellfäden erfolgt bei den meisten der Arten durch Teilung der terminal (=distal) sitzenden Zellen. Draparnaldia plumosa verfügt über morphologisch voneinander verschiedene Haupt- und Nebentriebe (= dicke und dünne Filamente). Die Zellen der Nebentriebe sind an ihren Oberflächen mit einem kohlenhydrathaltigen Belag versehen, der vermutlich Bakterien als Nahrungsquelle dient. Dünne Filamente sind, im Gegensatz zu den dicken, stets von ihnen umlagert.
Fritschiella ist eine Bodenalge, kommt aber auch in aquatischem Milieu vor. Je nach Habitat ändert sie ihre Wuchsform. Bodenbewohnende Arten bilden lange Fäden aus, von denen in den Luftraum hineinragende Nebentriebe abzweigen.
Oedogoniales: Die einreihigen, meist unverzweigten Zellfäden der Oedogoniales sind ausgesprochen polar gebaut. Das drückt sich bereits bei jeder interkalaren Zellteilung und dadurch aus, daß die Fäden eine basale Rhizoidzelle und apikale Spitzenzellen enthalten. Die Oedogoniales leben im Süßwasser, es sind meist festsitzende Vielzeller. Ihr Thallus ist fädig und unverzweigt. Hauptvertreter ist die Gattung Oedogonium. Deren Zellen enthalten einen netzförmig strukturierten, wandständigen Chloroplasten, sie sind einkernig. Asexuell pflanzen sie sich durch Zoosporen, sexuell durch Oogamie fort. Das Besondere an Oedogonium ist der außergewöhnliche Teilungsmodus der Zellen. Kernteilung und Zellteilung sind zeitlich synchronisiert, räumlich aber voneinander getrennt. Die Kernteilung findet, wie bei den meisten Pflanzenzellen üblich, nahe der Zellmitte statt. Während der Telophase wird zwischen den beiden Tochterkernen ein Septum, eine Vorstufe der späteren Querwand, angelegt, das zunächst noch ohne Kontakt zur Längswand ist. Während die Mitose vor sich geht, bildet sich nahe dem apikalen Pol - der Längswand innen ringförmig aufgelagert - ein Wulst aus plastischem (cellulosefreiem) Wandmaterial .
Im Anschluß an die Mitose werden der obere Tochterkern und das Septum apikalwärts verlagert. Dann reißt die Zellwand an einer präformierten Stelle außerhalb des Wulsts ringförmig auf. An seinem unteren Ende verwächst er mit dem Septum. Durch Auflagerung von weiterem Zellwandmaterial (Cellulose) differenzieren sich Septum und Wulst zu fertigen Quer- und Längswänden der neuen Tochterzelle. Die oberhalb der Sollbruchstelle verbleibenden Reste der Mutterzellwand bilden eine "Kappe" aus, die das typische Merkmal dieses Zellteilungsmodus ist. Am apikalen Pol aller interkalaren Zellen kann sich dieser Vorgang mehrfach wiederholen, so daß manche Zellen eine ganze Serie dieser Kappen tragen.
Die Arten sind diözisch, manche zeichnen sich durch einen ausgeprägten Sexualdimorphismus aus; dann sind die männlichen Pflanzen zu "Zwergmännchen" reduziert.
Die Ulvophyceen sind primär marine Organismen. Zoosporen sind, wenn vorhanden, symmetrisch gebaut. Sie besitzen zwei, vier oder mehr Geißeln. Ein Klassenmerkmal ist der deutlich ausgeprägte Generationswechsel mit haploiden Gametophyten und diploiden Sporophyten. Dauersporen werden nur ausnahmsweise gebildet. Den Ulvophyceae gehören folgende Ordnungen an:
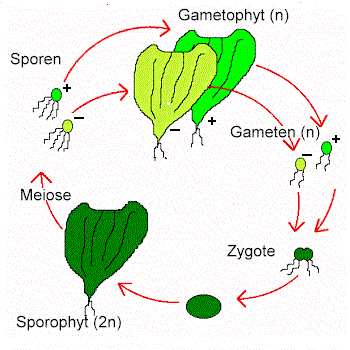
Codiolales: Die ehemals als Ordnung eingestuften "Ulotrichales" haben sich als heterogen erwiesen. Die Ulothrix ähnlichen Gattungen Uronema und Klebsormidium gehören anderen Klassen an. Enger umrissen sind die Codiolales. Charakteristisch in ihrem Lebenszyklus ist die große, photosynthetisierende Zygote, das "Codiolum"-Stadium, das eine Weile heranwachsen kann. In ihm findet die Meiose statt; es werden zweigeißlige Zoosporen frei, so daß sich die daraus entwickelnden Stadien haploid sind.
Der Ordnung gehören einzellige und mehrzellige Arten an, deren Thallus aus unverzweigten oder verzweigten Fäden besteht oder blattförmig (flächig) ist. Fast alle Arten leben benthisch. Sie sind durch nichtteilungsfähige Rhizoidzellen an Unterlagen verankert. Die meisten Arten und Gattungen (z.B. Acrosiphonia) sind marin, wenige leben im Süßwasser.
In einigen, doch nicht in allen Stammeslinien ist eine Progression von Isogamie zu Oogamie erkennbar (J. D. PICKETT-HEAPS, University of Colorado, Boulter, 1975).
Auffallend ist die jetzt in diese Ordnung einzubringende Gattung Ulothrix, von der die Art Ulothrix zonata am bekanntesten und häufigsten (im Süßwasser) ist. Ihr Thallus besteht aus unverzweigten Fäden (= + und - Gametophyt). Ihr Wachstum ist interkalar, und bei sexueller Fortpflanzung werden viergeißlige Zoosporen gebildet. Isogameten sind zweigeißlig. Sie entstehen nur unter Langtagbedingungen. Die Zygote hingegen keimt nur unter Kurztagbedingungen. Nach erfolgter Meiose zerfällt sie in vier bis acht Zoosporen, aus denen zur Hälfte + und zur Hälfte - Gametophyten herauswachsen. Die marine Art Ulothrix acorhiza vermehrt sich ausschließlich asexuell. Die Gattung Monostroma ist einschichtig flächig und leitet morphologisch zur nächsten Ordnung über.
 |
|
Ulvales: Ulva lactuca (Meersalat) und andere Ulva-Arten zeichnen sich durch einen flächigen, aus zwei Zellschichten bestehenden Thallus aus. Die Art ist ein Schulbeispiel für einen isomorphen heterophasischen Generationswechsel. Die Gameten sind getrenntgeschlechtig. Bei Enteromorpha ist der Thallus schlauchförmig, bei Prasiola stipitata wiederum flächig. Die Arten kommen im Süß-, Brack- und Salzwasser vor.
Cladophorales: Diese Ordnung hat eine Reihe eigenständiger Merkmale, die sich sowohl von den oben aufgeführten Ordnungen als auch von den folgenden Klassen abhebt, so daß auch die Ausgliederung zu einer neuen Klasse ("Cladophorophyceae") gerechtfertigt erscheinen mag. Hierzu gehören vielzellige Arten mit fädigem und (meist) verzweigtem Thallus. Das auffallende Merkmal ist die Vielkernigkeit der Zellen, denn Kern- und Zellteilung sind hier nicht synchronisiert. Die Fäden verlängern sich durch Spitzenwachstum, also durch Teilung apikal gelegener Scheitelzellen. Die Zellwand ist proteinreich, und als Gerüstkomponente enthält sie eine modifizierte Form der Cellulose (Cellulose I). Die Mikrofibrillen in übereinanderliegenden Schichten sind jeweils um 90 Grad gegeneinander versetzt, wodurch eine mikroskopisch gut sichtbare Schichtung zustande kommt. Die pyrenoidhaltigen Chloroplasten sind untereinander zu einem peripher gelegenen Netz verbunden. Haploide und diploide Phase sind, soweit vorhanden, isomorph, selten heteromorph. Die bekannteste Gattung ist Cladophora. Im marinen Bereich ist Cladophora vagabunda vorherrschend, im Süßwasser Cladophora glomerata. Letztere vermehrt sich ausschließlich vegetativ.
In dieser Klasse werden hier sämtliche Algen der "siphonalen Organisationstufe" zusammengefaßt (ohne Dasycladophyceae).
Bryopsidales: Hierher gehören durchweg einzellige Arten mit charakteristisch gestaltetem (siphonalem), vielfach verzweigtem, vielkernigem Thallus. Von wenigen Arten abgesehen (Bryopsis-, Derbesia- und Codium-Arten) sind sie ausschließlich in tropischen und sub-tropischen Zonen anzutreffen. Manche Arten zeichnen sich durch nur hier vorkommende Xanthophylle aus: Siphonein und Siphonoxanthin.
Bei Bryopsis ist der Thallus regelmäßig einfach oder vielfach einreihig verzweigt bis gefiedert; er kann bei den meisten Arten bis zu 10 Zentimeter groß werden, bei einer vor der japanischen Küste vorkommenden Art Bryopsis maximum erreichte er eine Größe von 40 Zentimetern. Bei der zweihäusigen Art Bryopsis plumosa sind männliche und weibliche Gametophyten getrennt; die männlichen Gameten sind kleiner als die weiblichen (Heterogamie, Anisogamie). Die Zygote wächst zu einem (einzelligen) verzweigten Faden heran. Es besteht noch Unklarheit über die Art und den Zeitpunkt der Meiose. Als ein Zwischenstadium erscheint nämlich ein "Riesenkern", der in eine Vielzahl kleiner Kerne zerfällt (meiotisch?, mitotisch?). Der Gametophyt entwickelt sich entweder direkt aus dem Sporophyten oder es wird ein Zoosporangienstadium zwischengeschaltet. Die nah verwandte Art Bryopsis hypnoides ist einhäusig. In der Gametophytenzellwand kommen Cellulose und Xylan vor, die Wand des Sporophyten besteht vornehmlich aus Mannan.
Der wenige cm große Sporophyt von Derbesia marina ähnelt Bryopsis. Er ist siphonal, verzweigt, einreihig. Gelegentlich sind ältere Thallusabschnitte durch Querwände abgegliedert. Der zugehörige Gametophyt ist ganz anders gestaltet und unter dem Namen Halicystis ovalis bekannt. Die ca. 1 cm großen Bläschen sind kugelförmig und sitzen mit rhizoidartigen Verzweigungen fest. Wir haben es hier wider mit einem typischen Beispiel für einen heteromorphen, heterophasischen Generationswechsel zu tun. Die Gameten sind unterschiedlich groß (anisogam). Wie bei Bryopsis unterscheiden sich die Thalli beider Phasen in ihrer Zellwandzusammensetzung.
Codiales: Manche Arten kommen in der Nordsee vor. Die schon makroskopisch auffälligen, mehrere Dezimeter großen, oft gegabelten oder kugelförmigen Thalli dieser Ordnung bestehen aus vielfach verzweigten siphonalen Zellfäden, die zu einem Geflecht verwoben sind. Bei Codium besteht die Oberfläche aus einer Schicht keulenförmiger Ausstülpungen, die wie Palisadenzellen der Blätter höherer Pflanzen plastidenreich sind. An der Basis entwickeln sie zu Zeiten die kleineren, keulenförmigen Gametangien, in denen die Meiose stattfinden soll. Die Wand besteht aus einem beta-1,4 -Mannangerüst, das in Arabinogalactan eingelagert ist.
Caulerpales: Bei Caulerpa sitzen an langen Ausläufern einerseits Rhizoidbündel, andererseits entspringen von dort blattartige, aufrechte Teile. Diese bis zu mehrere Dezimeter groß sein. Die Festigkeit der Struktur wird durch eingezogene Querbalken aus Wandmaterial erhöht, deren Gerüstsubstanz beta-1,4 -Xylan ist, welchem beta-1,3 -Glucan assoziiert ist. Bemerkenswert ist, daß hier neben den Chloroplasten auch Leukoplasten vorkommen. Die Pflanzen sind diploide Gametophyten.
Für die enge Verwandtschaft mit den Bryopsidophyceae sprechen der siphonale Bau der Thalli, die chemische Zusammensetzung der Zellwände, vor allem aber die Unterschiede zwischen Gametophytenwand und der der Cysten (Sporophytenwand).. Im Gegensatz zu den Caulerpales sind die Zellen lange Zeit einkernig. Der Thallus ist radiärsymmetrisch gebaut. Die Fortpflanzung erfolgt über Cysten, aus denen Isogameten entlassen werden.
Zu den Dasycladiales gehört Acetabularia, ein Standardobjekt biologischer Grundlagenforschung. Diese Gattung ist mit mehreren Arten an den Meeresküsten wärmerer Zonen vertreten. Acetabularia mediterranea ist im Mittelmeer häufig, Acetabularia crenulata in der Karibik. Der Thallus ist an natürlichen Standorten meist verkalkt. Im Labor hingegen werden Kulturbedingungen gewählt, die eine Verkalkung unterbinden. Eine Zelle ist in die drei Abschnitte Rhizoid, Stiel und Hut untergliedert .
Der Zellkern (Primärkern) liegt im Rhizoid. Er enthält u.a. Lampenbürstenchromosomen (H. SPRING et al., 1975) und ist vermutlich polyploid. Vor Beginn der Cysten- und Gametenbildung wandert er unter meiotischer und mitotischer Aufteilung in 10-15 000 Sekundärkerne durch den Stiel in Richtung Hut. Die unterwegs entstandenen Produkte verteilen sich auf dessen Septen.
Zygnematales: Zygnematales, auch Konjugaten genannt, sind durch eine Variante ihres Fortpflanzungsverhaltens, die Konjugation, gekennzeichnet. Nie werden begeißelte Gameten gebildet. Die Protoplasten der Konjugationspartner nehmen vorübergehend amöboide Gestalt an. Die aus der Gametenverschmelzung (Fusion) hervorgehende Zygote umgibt sich mit einer dicken Wand und ist in der Regel als ein Dauerstadium (Hypnozygote) anzusehen. Nach langer Reifezeit, oft im Jahreszyklus, werden haploide Zellen entlassen, aus denen sich, je nach Art, einzellige oder fädige Gametophyten entwickeln. Die Zygnematales unterteilt man üblicherweise in drei Familien: Zygnemataceae, Mesotaeniaceae, Closteriaceae und Desmidiaceae.
Den Zygnemataceae gehören die im Süßwasser verbreiteten fädigen Spirogyra-, Mougeotia- und Zygnema-Arten an. Die Gattungen unterscheiden sichdurch die Struktur ihrer Chloroplasten. Spirogyra enthält einen schraubig gewundenen, bei Mougeotia ist er plattenförmig und um seine Längsachse drehbar. Unter Starklichtbedingungen orientiert er sich mit der Kantenseite zur Lichtquelle, unter Schwachlichtbedingungen mit der Breitseite. Zygnema- und Zygnemopsis-Arten besitzen sternförmige Chloroplasten. Zur Einleitung der Konjugation werden zwischen Zellen benachbart liegender Fäden Brücken ausgebildet, durch die der Protoplast der Zelle des einen Fadens in den benachbarten überwechselt.
Die Mesotaeniaceae sind einzellig. Die Zellen sind von einer einheitlichen, in sich geschlossenen Zellwand umgeben. Die asexuelle Fortpflanzung erfolgt durch Querteilung der meist länglichen Zellen. Vor der Konjugation formt jede der an der Paarung beteiligten Zellen Ausstülpungen (Papillen), die zu einer gemeinsamen brückenähnlichen Verbindung verschmelzen, innerhalb derer die Protoplasten fusionieren und eine Zygote bilden. Die bekanntesten Gattungen sind Mesotaenium, Spirotaenia, Cylindrocystis, Netrium. Nah verwandt mit den Mesotaeniaceae sind die Closteriaceae.
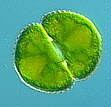 Die
Desmidiaceen unterscheiden sich von den beiden übrigen Familien primär
durch den Mechanismus ihrer vegetativen Fortpflanzung und die dadurch bedingten
Eigenarten in der Architektur der Zellwand. Die Zellen bestehen aus zwei
Halbzellen, von denen eine jünger als die andere ist. Zwischen ihnen
liegt, eingeengt durch eine mehr oder weniger tiefe Einschnürung (=Sinus),
der Isthmus. Die Wand ist nicht durchgehend, vielmehr besteht sie aus zwei
einander überlappenden Halbschalen.
Die
Desmidiaceen unterscheiden sich von den beiden übrigen Familien primär
durch den Mechanismus ihrer vegetativen Fortpflanzung und die dadurch bedingten
Eigenarten in der Architektur der Zellwand. Die Zellen bestehen aus zwei
Halbzellen, von denen eine jünger als die andere ist. Zwischen ihnen
liegt, eingeengt durch eine mehr oder weniger tiefe Einschnürung (=Sinus),
der Isthmus. Die Wand ist nicht durchgehend, vielmehr besteht sie aus zwei
einander überlappenden Halbschalen.
 Nach
der eigentlichen Zellteilung durch ein irisblendenartig eingezogenes Septum
im Isthmus stülpt jede Halbzelle eine neue Tochterhalbzelle aus. Während
diese heranwachsen und sich differenzieren, rücken die beiden alten
Halbzellen auseinander. Die Tochterzellen trennen sich, nachdem sie nahezu
ihre volle Größe erreicht haben. Die Entwicklungsstadien sind
bei einer Reihe von Arten studiert und durch eindrucksvolle Bildserien
dokumentiert worden.
Nach
der eigentlichen Zellteilung durch ein irisblendenartig eingezogenes Septum
im Isthmus stülpt jede Halbzelle eine neue Tochterhalbzelle aus. Während
diese heranwachsen und sich differenzieren, rücken die beiden alten
Halbzellen auseinander. Die Tochterzellen trennen sich, nachdem sie nahezu
ihre volle Größe erreicht haben. Die Entwicklungsstadien sind
bei einer Reihe von Arten studiert und durch eindrucksvolle Bildserien
dokumentiert worden.
Wachsende Tochterhalbzellen sind von einer Primärwand umgeben,
die im Verlauf des weiteren Wachstums durch eine darunter angelegte Sekundärwand
abgelöst wird. Nach deren Fertigstellung wird die Primärwand
entweder abgestoßen (wie z.B. bei Cosmarium), oder sie löst
sich sukzessive auf (z.B. bei Cosmocladium saxonicum). Bei der zuletzt
genannten Art sind an den Oberflächen von Primär- und Sekundärwand
unterschiedliche Glykokonjugate exponiert. Neben speziellen Bewegungsgallerten
sind verschiedentlich Konjugationsgallerten (Kopulationsgallerten) nachgewiesen
worden, die sich während der Papillenbildung, d.h. während früher
Stadien des Konjugationsprozesses bilden.
Die Wände der Desmidiaceen sind vielfach durch Auflagen verstärkt, mit Ornamenten versehen, und von regelmäßig angeordneten Poren durchbrochen. Diese enthalten spezifisch strukturierte Gallertpfropfen (Porenkomplexe), die sich bereits lichtmikroskopisch durch einschlägige Färbungen (z.B. mit Methylenblau) aber auch elektronenmikroskopisch oder durch Einsatz fluoreszenzmarkierter Lektine identifizieren lassen
Von Ausnahmen abgesehen (Staurastrum u.a.), gehören die
Arten dieser Familie nicht zu den häufigen, wegen ihrer symmetrischen
Zellformen aber zu den auffälligsten Algen. Sie bevorzugen Biotope
mit niedrigem pH-Wert (pH 4-5).
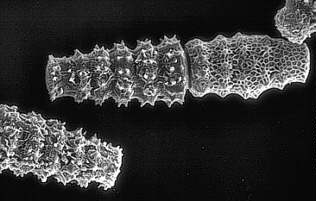
Pleurotaenium kayei (Aufn.: M. ENGELS). Diese Aufnahme wie auch auch alle übrigen Zygnematales-Bilder in Botanik online wurden von Kulturen aus der Sammlung von Conjugaten-Kulturen (SVCK) am Institut für Allgemeine Botanik der Universität Hamburg gemacht.
Arten der Gattungen Micrasterias, Cosmarium, Euastrum, Staurastrum, Pleurotaenium
u.a. sind einzellig, die der Gattungen Hyalotheca, Desmidium, Spharozosma u.a. fädig. Teilungen erfolgen auf die gleiche Weise wie bei den einzelligen Arten. Wir haben es daher mit einem interkalaren Wachstum zu tun.
Die Bildung des Septums zeigt streckenweise Anklänge an die Bildung von Zellwänden in einem Phragmoplasten. Letzteres ist aber charakteristisch für die Zellteilung in der folgenden Klasse. Moderne Autoren fassen daher die Zygnematophyceae gelegentlich mit ihr zusammen oder ordnen sie ihr ein.
Charyophyceae im traditionellen Sinne sind die Armleuchteralgen mit Chara und Nitella als repräsentativen Vertretern. Die Zell- und Kernteilung ähnelt der der höheren Pflanzen; die äußere Gestalt dieser Algen weist auch in diese Richtung. Dennoch weiß man schon seit langem, daß die Armleuchteralgen nicht als Vorfahren der Landpflanzen in Betracht kommen, weil sie in zu vielen anderen Merkmalen als zu spezialisiert betrachtet werden können.
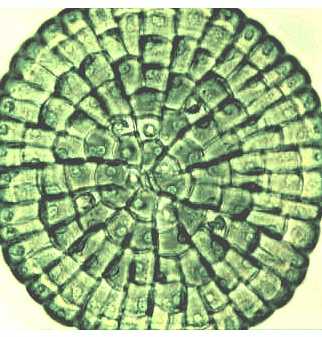 Choleochaete repräsentiert eine rezente Grünalgengruppe (den Charophyceae angehörend), deren Vorfahren mit den frühesten Landpflanzen verwandt waren. Da nur wenige fossile Funde zum Übergang Algen - Landpflanzen vorliegen, bedarf es des genauen Studiums der rezenten Arten, um den Evolutionsprozeß der zur Landpflanzenentstehung führte, rekonstruieren zu können - Aufn.: Linda E. GRAHAM - © Botany for the Next Millenium |
Am interessantesten ist jedoch die Gattung Coleochaete (Ordnung: Coleochaetales), die früher stets zu den Ulotrichales gestellt wurde, sich aber durch einen Kern- und Zellteilungsmechanismus auszeichnet, der dem der Landpflanzen gleicht. Hinzu kommt, daß in den Peroxysomen von Coleochaete das Enzym Glycolatoxydase nachgewiesen wurde, das ebenfalls für höhere Pflanzen typisch ist. Schließlich sind auch noch in geologischen Formationen, in denen die ersten Landpflanzen gefunden wurden, Coleochaete-ähnliche Fossilien nachgewiesen worden (L. E. GRAHAM, 1984). Die nächsten Jahre werden sicher weitere Belege zutage fördern, die darüber entscheiden, ob Coleochaete-ähnliche Formen den Vorstufen von Landpflanzen nahekommen oder ob andere Kandidaten hierfür gesucht werden müssen. Nach anderen Ansichten sollte man Coleochaete einer eigenen Ordnung zuweisen und sie dann gemeinsam mit den Klebsormidiales zu den "Klebsormidiophyceae" stellen.
Charales: Sie besitzen einen hoch organisierten Thallus, der in Knoten (Nodien) und Internodien untergliedert ist. An den Nodien sitzen in quirliger Anordnung Kurztriebe ("Blätter"), deren Zellen nur begrenzt teilungsfähig sind. In den "Blattachseln", können unbegrenzt weiterwachsende Langtriebe abzweigen. Chara-Arten kommen auf sandigen Böden am Grunde klarer stehender Gewässer vor. Durch Rhizoide (Studienobjekte der Geotropismusforschung) sind sie im Boden verankert. Chara bevorzugt alkalisches Milieu, man findet sie aber auch im Brackwasser; sie ist jedoch phosphatempfindlich und kommt deshalb in eutrophen Gewässern kaum mehr vor, ein Grund, weshalb sie in unseren Breiten sehr selten geworden ist.
Das Wachstum der Achsen erfolgt mit einschneidigen Scheitelzellen; die Differenzierung in die einzelnen Abschnitte beruht auf einer strikten Zellteilungsfolge. Die von der Scheitelzelle abgegliederte Zelle teilt sich zunächst inäqual in eine Nodien- und eine Internodienzelle. Aus der Nodienzelle entwickeln sich parenchymatische Zellkomplexe, aus deren Peripherie die Scheitelzellen der Kurztriebe entstehen. Die Internodiumzelle wächst unter Kernteilungen enorm heran. Sie bildet eine große Zentralvakuole und enthält in ihrem plasmatischen Wandbelag schließlich eine hohe Zahl von Chloroplasten und bis über 1000 Zellkerne. Ausgewachsene Internodien können sehr groß (bis zu 10 cm) sein, ihre Länge entspricht jeweils einer einzigen Zelle. Das Zellplasma ist zweiphasig.
Die äußere chloroplastenhaltige Phase ist stationär, die innere in ständiger Bewegung. Die leicht zu beobachtende "Rotationsströmung" zieht in starker Steigung schraubenförmig auf einer Zellseite nach oben, auf der anderen nach unten. Zellteilung zu Zellteilung wechselnde Orientierung der Kernspindel führt zur Ausbildung regelmäßiger Muster. "Sprosse" und Rhizoide sind in erheblichem Maße regenerationsfähig. Die Zellen der Charophyceen können sehr groß sein. Die Länge der Internodien entspricht der Länge einzelner Zellen. Bei den Chara-Arten sind sie von kürzeren, parallel angeordneten Rindenzellen umgeben.
Chara- und Nitella-Arten sind daher als Versuchsobjekte zum Studium intrazellulärer Bewegungen herangezogen worden. Nitella eignet sich darüber hinaus als Modell für elektrophysiologische Untersuchungen pflanzlicher Zellen. Die Zellen sind groß genug, um eine Glaselektrode in sie einzuführen und das Membranpotential abzuleiten.
Die Internodien der Chara-Arten sind berindet. Parallele Fäden bedecken die Zentralzelle lükkenlos. Diese Berindung geht von Initialen der Nodien aus und wächst mit der Streckung der zentralen Nodienzelle mit. Die die Oberfläche bedeckenden Zellfäden folgen dabei dem gleichen strengen Differenzierungsschema wie die Zellen der Hauptachse. Das gilt auch für die Rhizoide. Achen und Rhizoide sind in erheblichem Maße regenerationsfähig. Bei der Gattung Nitella fehlt die Berindung.
Außer dem charakteristischen Bau der vegetativen Teile zeichnen sich Charophyceen durch komplex gebaute Fortpflanzungsorgane aus. Es gibt monözische und diözische Chara-Arten. der Befruchtungsmodus ist die Oogamie. Ungewöhnlich für Algen sind die einzeln stehenden Eizellen, die von schraubig angeordneten sterilen Hüllzellen umgeben sind. Auch die sogenannten spermatogenen Fäden, aus denen sich die männlichen Gameten entwickeln, sind von spezialisierten, plattenförmigen, zu einer Hohlkugel zusammenschließenden, sterilen Zellen umgeben. Die zweigeißligen Spermatozoiden sind schraubig verdreht und erinnern an die Torfmoose.
Charophyceenthalli sind oft verkalkt und daher auch fossilienbildend. Die Struktur der Oogonienhülle erwies sich dabei als wichtiges Erkennungszeichen. Versteinerte Formen kommen seit dem Devon vor.
|
|