
Wie beschrieben , ist der Vegetationskörper aller höheren Pflanzen von einem Leitbündelsystem durchzogen. Es untergliedert sich in Xylem und Phloem, das von Festigungsgewebe umgeben sein kann. Im allgemeinen gilt, daß die Wasser- und Ionenversorgung über das Xylem, der Assimilattransport - und die Verteilung weiterer Substanzen - über das Phloem erfolgt. Innerhalb des Xylems ist der Transport acropetal, d.h., zur Sproßspitze hin gerichtet. Die Mobilisierung der Substanzen im Phloem kann basipetal, d. h. in Richtung Wurzel, oder bidirektional (acri- und basipetal) erfolgen.
Das Xylem lebender Pflanzen (auch als Holz- oder Gefäßteil bezeichnet) ist ein zusammenhängendes, wasserhaltiges apoplastisches System abgestorbener Zellen von beträchtlicher Ausdehnung (ein System kommunizierender Röhren). Es dient dem Transport des Wassers und darin gelöster Ionen. Der Gefäßteil reicht vom Blatt über den Sproß bis zur Wurzel, wobei er nicht direkt an die Außenwelt, sondern an lebende Zellen grenzt. Selbst wenn sich die Landpflanzen völlig gegen den Verlust von Feuchtigkeit schützen, also die Transpiration verhindern könnten (was jedoch nicht möglich ist), müßten sie doch Wasser für ihre Wachstumsvorgänge aufnehmen. Der Hauptteil des für Transpiration und Wachstum benötigten Wassers wird aus der Bodenfeuchtigkeit über das Wurzelsystem aufgenommen.
Allgemein anerkannt ist heute die Tatsache, daß Pflanzen für den Langstreckentransport der Feuchtigkeit nicht aktiv (aus der Photosynthese stammende) Energie aufbringen müssen, sondern daß er passiv erfolgt. Aktive Vorgänge (Ionenaufnahme) dagegen rufen einen Wurzeldruck hervor. Die Energie für den Langstreckentransport stammt letzlich, wie die zur Photosynthese benötigte Energie, aus der Sonnenstrahlung.
Der Übergang von der wässrigen in die Dampfphase beginnt in den Atemhöhlen der Blätter. Hier verdunstet Wasser von den Oberflächen der an die Atemhöhlen grenzenden Zellen. Die Energie dazu stammt entweder direkt aus der Sonnenstrahlung, oder aus der Wärmeenergie der die Pflanze umgebenden Atmosphäre.
Nach welchem Förderprinzip wird nun das Wasser bis in die Gipfel der höchsten Bäume transportiert? Der meist vor Blattaustrieb bei manchen Baumarten deutlich ausgeprägte Wurzeldruck, reicht dazu ganzjährig nicht aus. Nach der Kohäsionstheorie des Wassertransports, sollen: " .. an den verdunstenden Blattzellen .." Wasserfäden im Xylem hängen: " .. deren untere Enden mit dem Bodenwasser in Verbindung stehen" (nach J. BÖHM [1831 - 1893], Hochschule f. Bodenkultur, Wien). Im Verlaufe der Transpiration entwickle sich als Folge des Wasserverlusts ein Sog auf diese, durch Kohäsionskräfte zusammengehaltenen, hängenden Wasserfäden. Genügend Bodenfeuchtigkeit vorausgesetzt, entstehe dann eine gezogene Volumenströmung, der sog. Transpirationsstrom, der den Feuchtigkeitsverlust der Blätter ausgleicht. An der Theorie wurden immer wieder Zweifel laut und alternative Modelle sind entwickelt werden.
Wasseraufnahme durch die Wurzel: Die Absorption erfolgt durch die, in direktem Kontakt mit dem Boden stehende, Rhizodermis. An den jüngsten Wurzelspitzen wird ihre wasser- und ionenaufnehmende Fläche durch die Ausbildung von Wurzelhaaren vergrößert. Von hier aus bewegt sich die Bodenlösung weiter durch das Rindenparenchym. Schließlich erreicht sie die innerste Rindenschicht, die Endodermis. Diese umhüllt den Zentralzylinder, der das Phloem und das Xylem enthält. Vom Xylem wird das Wasser mit den darin gelösten Ionen aufgenommen und dieser Xylemsaft weiter in Richtung Sproßspitze transportiert. In älteren Abschnitten der Wurzel, in denen die Rhizodermis schon abgestorben ist, wird die Wurzel nach außen von der ein- bis mehrschichtigen Exodermis begrenzt. Die Wände ihrer Zellen sind verkorkt.
Die Passage der einzelnen Schichten wird in entscheidendem Maße von ihrer jeweiligen Struktur bestimmt. Jedes der Wurzelgewebe kann im Prinzip auf drei verschiedene Arten vom Wasser durchquert werden.
- apoplastisch, d. h. durch die Zellwände und die Interzellularen
- symplastisch, d. h. von Protoplast zu Protoplast via Plasmodesmen
- transzellulär, von Zelle zu Zelle, wobei auch die Vakuolen passiert werden.
Alle drei Wege sind außerhalb der Endodermis möglich. Hier angelangt, scheidet für die Bodenlösung der apoplastische Weg aus. Er wird durch eine wasserabweisende Inkrustation in der Zellwand, den Casparischen Streifen, wirkungsvoll behindert. Diese Sperre ermöglicht der Pflanze eine selektive Ionenaufnahme. Es gibt auch Pflanzen, bei denen der Casparische Streifen bereits in der Exodermis liegt, so daß die apoplastische Barriere schon hier zur Geltung kommt
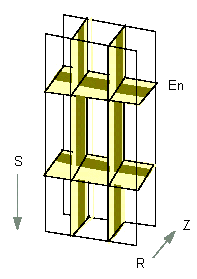
Dreidimensionales Schema von Zellen der Wurzelendodermis (Längsschnitt). Der Casparische Streifen ist dunkeloliv hervorgehoben (En: Endodermis, S: Richtung Wurzelspitze, Z: Zentralzylinder, R: Rinde (Nach U. LÜTTGE und N. HIGINBOTHAM, 1979).
Noch nicht vollständig geklärt ist das Transportprinzip des Wassers beim Durchtritt durch die Endodermis. Nach der Kohäsionstheorie, die eine gezogene Volumenströmung annimmt (siehe oben), entspräche die Wasserbewegung durch die gesamte Wurzel einer Unterdruckfiltration. Die Energie für diese (passive) Art der Feuchtigkeitsbewegung stammt aus einem Gefälle des Wasserpotentials zwischen dem Boden, dessen Wert meist hoch liegt, und dem niedrigerem der Wurzel. Genanntes Potential stellt eine volumenbezogene Energieform [J/m3] dar. Als solche kann es auch in Druckeinheiten (Pascal oder Bar) angegeben werden. Das Wasserpotential ist definiert als der negative Wert des Drucks, mit dem ein System Wasser aus dem Bezugssystem aufnimmt. Als Bezugssystem gilt reines Wasser, das sich bei einer Temperatur von 25 �C auf Meereshöhe und unter dem atmosphärischen Normaldruck von 1,01325 bar befindet. Das Potentialgefälle wird durch direkte Sonnenstrahlung und durch das permanente Wasserdampfdefizit der Atmosphäre aufrecht erhalten. Selbst bei niedrigem Defizit, also hoher Luftfeuchtigkeit, sind doch die aufnahmefähigen Luftmassen enorm. Sie verringern laufend den Inhalt der pflanzlichen Feuchtigkeitsspeicher. Die Anwesenheit eines deutlichen hydrostatischen Drucks in der Wurzel, kann von der Kohäsionstheorie nicht erklärt werden. Hier sind aktive, energieverbrauchende Vorgänge wirksam. Der Druckaufbau erfolgt über osmotische Vorgänge.
Wasserabgabe durch das Blatt: Neben der dampfförmigen Abgabe von Wasser im Verlaufe der Transpiration, kann an manchen Pflanzen ein Abtropfen flüssigen Wassers beobachtet werden (Guttation). In der Flüssigkeit können auch Salze gelöst sein. Diese Pflanzen verfügen an den Blatträndern über (aktive oder passive) Hydathoden (Wasserspalten) oder auch über Stomata, wo das Wasser (bzw. die Lösung) in feinen Tropfen abgesondert werden kann.
Es ist erwiesen, daß im Phloem Assimilate, Phytohormone, Aminosäuren, Herbizide, Viren u.a. transportiert werden. Assimilate werden u.a. zu den Speicherorganen der Wurzel, der Früchte und der Samen befördert. Für Phytohormone wurde ein acropetaler und ein basipetaler Transport nachgewiesen. Für manche Viren wurde gezeigt, daß sie sich über die gesamte Pflanze ausbreiten. Im Phloem wurden Transportgeschwindigkeiten von 30-150 cm/Stunde gemessen, was bedeutet, daß eine 0,5 mm langes Siebelement bei einer durchschnittlichen Durchflußgeschwindigkeit von 90 cm/Stunde alle zwei Sekunden neu beladen wird.
Der Einsatz von radioaktiv markierten Substanzen und das Verfahren der Autoradiographie erlaubten es, den Stofftransport (Translokation) in pflanzlichen Geweben zu verfolgen. Während der Xylemtransport durch externe Energiezufuhr (Wärme, Wasserpotential) relativ leicht erklärbar ist, ist die Frage nach der treibenden Kraft im Phloem weniger offensichtlich. Das Problem ist in zwei Teile zu untergliedern:
- Phloembeladung
- Phloemtransport
Die Phloembeladung, also das Beschicken der Siebzellen in den Blättern mit Transportgut, ist selektiv und energieabhängig (ATP-Verbrauch); es sind offensichtlich membrangebundene Pumpen am Werk. Saccharose reichert sich im Phloem schneller an als andere Zucker, bei einigen Pflanzenarten werden bestimmte Aminosäuren ins Phloem gepumpt. Hinzu kommen einige Ionen (Stickstoff, Phosphor, Kalium [jedoch nicht Calcium, Eisen und Bor]), ferner Herbizide (2,4,5 T, 2,4 D.) sowie Phytohormone. Durch den aktiven Transport bedingt, baut sich in den Siebelementen ein osmotischer Druck auf, der etwa doppelt so hoch wie in den Zellen der Umgebung (Zellen der Gefäßbündelscheide) ist. Für die große Differenz sind in erster Linie die Zucker verantwortlich.

E. MÜNCH (Forstbotanisches Institut, Universität München, 1926) postulierte, daß der sich hier aufbauende Druck die treibende Kraft der Strömung im Phloem ist. Für die Mobilität wäre demnach nicht der Inhalt, sondern die Effizienz der Beladung entscheidend. Nach MÜNCHs Annahme (Druckstrom-Hypothese) müßte sich der Phloeminhalt passiv bewegen, eine Energiezufuhr unterwegs wäre demnach nicht erforderlich. Die Energie resultiert aus dem Druckunterschied, und es baut sich daher ein Gradienten zwischen Beladungs- und Entladungsort auf. Wie am Beispiel morphogener Gradienten beschrieben, würde es sich dabei jedoch um ein sehr labiles System handeln. Zudem könnte der Transport nur in eine Richtung erfolgen. Dem steht die Feststellung entgegen, daß es in der Tat einen bidirektionalen Transport gibt (W. ESCHRICH, Forstbotanisches Institut Göttingen, 1970) und daß das Transportgut unterschiedlich schnell verteilt wird. Substanzen aus jungen Blättern werden meist basipetal, die aus älteren Blättern oft acropetal befördert. Im Stengel überschneiden sich die Wege. Allein hierdurch würde es zu einem Druckausgleich kommen, wodurch der Transport zum Erliegen käme.
|
|