
Der charakteristische Biosyntheseprozeß grüner Pflanzen ist die Biosynthese von Kohlenhydraten via Photosynthese. Wegen der Bedeutung und der Komplexität der dabei ablaufenden Prozesse werden sie in einem gesonderten Abschnitt behandelt.
An dieser Stelle ist es nur wichtig zu wissen, daß dabei Glucose und andere (phosphorylierte) Zucker entstehen. Wie wir gesehen haben, wird Glucose keineswegs nur abgebaut, sondern ist Ausgangsprodukt z.B. von Biosynthesewegen vieler Polysaccharide, die einerseits als Strukturelemente der Pflanzenzelle (z.B. Cellulose als Wandkomponente), zum anderen als Speicherstoffe (z.B. Stärke) dienen. Darüber hinaus enthalten Pflanzenzellen ein weites Spektrum verschiedener, doch strukturell sehr ähnlicher Zucker. Eine wichtige Frage lautet nun: Wie werden sie ineinander überführt, und was sichert die Selektivität der einzelnen Reaktionsschritte?
Wenn nicht schon geschehen, müssen die Zucker, wie alle übrigen Ausgangssubstanzen von Biosyntheseprozessen, zunächst aktiviert werden. Dies geschieht hier entweder durch Übernahme von Phosphatgruppen oder durch Bindung des Zuckerrests an ein Nukleotid.
Fructose-6-Phosphat ist ein wichtiges Intermediärprodukt der Photosynthese und der Glykolyse. Es steht mit dem Glucose-6-Phosphat, und dieses wiederum mit dem Glucose-1-Phosphat, im Gleichgewicht.
Glucose-1-Phosphat und UTP ergibt UDP-Glucose. UDP-Glucose + Fructose-6-Phosphat polymerisieren unter UDP-Abspaltung zu Saccharosephosphat, welches unter Phosphatverlust in Saccharose (Rohrzucker, ein Disaccharid) überführt wird. Die Reaktion findet in Chloroplasten statt.
Der Weg zur Stärke führt über Glucose-1-Phosphat und dessen Kopplung an Adenosindiphosphat
Glucose-1-Phosphat + ATP + H2O > ADP-Glucose + 2 Pi
Demnach entscheidet die Bindung an ein Nukleosiddiphosphat, welcher Reaktionsweg eingeschlagen wird.
UDP-Glucose (nicht aber ADP-Glucose) kann auch in Glykolipide und Glykoproteine eingebaut werden.
Zur Stärkebildung werden zwei Enzyme benötigt, eines für den Start und die Kettenverlängerung (alpha 1 > 4 glykosidische Bindung) durch Monosaccharideinheiten, das zweite zum Einfügen der alpha 1 > 6 glykosidischen Bindungen sowie der Verknüpfung von Ketten untereinander (über alpha 1 > 4 glykosidische Bindungen). Dieses Enzym koppelt nicht einzelne Glucosyleinheiten an vorhandene Ketten, sondern es werden gleich ganze Ketten übertragen. Man unterscheidet dabei zwischen intramolekularem und intermolekularem Transfer.
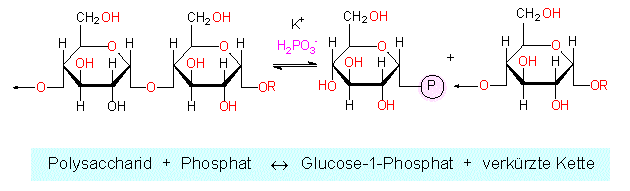
Für den Stärkeabbau werden in Pflanzenzellen mindestens vier Enzyme benötigt. Es gibt zwei alternative Wege:
Hydrolytischer Abbau durch eine Amylase. Hierbei wird die Stärke in Disaccharideinheiten (Maltose) zerlegt. Ein Abbau in Nähe von Verzweigungspunkten ist nicht möglich. Es verbleibt daher ein Rest: das Grenzdextrin.
Phosphorylytischer Abbau durch eine Phosphorylase. Hierzu ist anorganisches Phosphat unerläßlich. Das Endprodukt ist Glucose-1-Phosphat Die Reaktion ist im Gegensatz zur Amylasereaktion reversibel.
Umwandlung eines Zuckers in einen anderen durch Epimerisierung:
Ein Beispiel hierfür ist die Umwandlung von Glucose in Galactose. Ausgangspunkt ist wieder ein an UDP gebundener Glucosylrest. NAD+ wird benötigt, doch kommt es am Ende der Reaktion unverbraucht wieder heraus).
UDP-D-Glucose + NAD+ > UDP-4-Keto-D-Glucose + NADH + H+
UDP-4-Keto-D-Glucose + NADH + H+ > UDP-D-Galactose + NAD+
Bildung von C5-Zuckern: Der oxydative Pentosephosphatzyklus: Der oxydative Pentosephosphatzyklus ist in vielem die Umkehr des reduktiven Pentosephosphatzyklus (= CALVIN-Zyklus), den wir später im Detail im Zusammenhang mit der Photosynthese kennenlernen werden.
Im Gegensatz zum CALVIN-Zyklus, der in den Chloroplasten abläuft, finden die Reaktionen des oxydativen Pentosephosphatzyklus im Cytosol statt; zudem sind die Reaktionen nicht auf Pflanzenzellen beschränkt. Durch Oxydation von Glucose-6-Phosphat und Kohlendioxyd-Abspaltung entsteht Ribose-5-Phosphat, wobei Elektronen vom NADP+ übernommen werden.
Glucose-6-Phosphat + 2 NADP + H2O > Ribose-5-Phosphat + 2 NADPH + H+ + CO2.
Die Bedeutung von Ribose-5-Phosphat braucht eigentlich nicht extra erwähnt zu werden, denn es ist ein wesentlicher Bestandteil von ATP, CoA, NAD+, FAD, RNS, DNS u.a.
Zu den weiteren Reaktionen gehören Umwandlungen zu C3-, C4-, anderen C5- und C6-, sowie einem C7-Zucker. Dabei sind sowohl Transketolasen als auch Transaldolasen beteiligt.
Einige der Reaktionen führen zu Intermediärprodukten, die auch im Verlauf der Glykolyse auftreten. Sie stellen damit Bindeglieder zwischen dem Pentosephosphatzyklus und der Glykolyse dar. So entsteht z.B. aus zwei (phosphorylierten) Fünfer-Zuckern (Xylulose-5-Phosphat und Ribose-5-Phosphat) das Glycerinaldehyd-3-Phosphat und ein Siebener-Zuckerderivat, das Sedoheptulose-7-Phosphat. Dabei wird eine Ketogruppe von einem auf den anderen Zucker übertragen, während als Beiprodukt ein Intermediärprodukt der Glykolyse entsteht.
Andererseits können die beiden Produkte in einer Transaldolasereaktion zurückreagieren, wobei Erythrose-4-Phosphat und Fructose-6-Phosphat gebildet werden. Letzteres ist ein weiteres Zwischenprodukt der Glykolyse; ersteres wird, wie wir schon gesehen haben, für die Synthese der Aminosäuren Serin und Glycin benötigt.
Welcher der einzelnen Wege eingeschlagen wird, hängt zu einem überwiegenden Teil von den Konzentrationen an ATP, NAD+, NADP+ u.a. ab. Ihr Bedarf und Angebot entscheidet darüber, ob ein Intermediärprodukt im Pentosephosphatzyklus verbleibt oder über die Glykolyse abgebaut wird. Andererseits eröffnet sich durch die Kopplung beider Wege eine Möglichkeit zur Glucoserückgewinnung. Wir haben bei der Besprechung der Glykolyse gesehen, daß zwei der Schritte irreversibel sind, d.h. Glucose kann über eine Umkehr der Glykolyse nicht zurückgebildet werden, wohl aber, wenn ein Intermediärprodukt (Glycerinaldehyd-3-Phosphat) in den oxydativen Pentosephosphatzyklus eingespeist wird und das Gleichgewicht dort auf der Seite der Glucose-Resynthese liegt.
|
|
 Ribose-5-Phosphat
Ribose-5-Phosphat 


