

Ability of a recessive (upper part) and a dominant (lower part) allele to assert itself under the precondition that it has a selective advantage of 1 in 1,000 (s = 0.001). The ordinate gives the allele frequency between 0 and 1, the abscissa the number of generations from 1 to 10,000 (according to T.O.SOLBRIG, 1970).
How large is the probability that a mutation asserts itself in a sexually propagating population? Neglecting selection, R.A. FISHER could show that a once arisen mutation symbolized by the genotype Aa in a population of individuals with the genotype AA has hardly any chance to survive. In order to hand the allele a down to its offspring, the Aa individual has to pair with an AA individual. The probability of a loss of the allele a is given by FISHER as being e –1 per generation. The rate of elimination is described by a Poisson-distribution:
e-1 = 0,368.
The probability that the allele a does still exist after one generation is therefore
1 - 0,368 = 0,632
and after another generation:
e-(1- 0368) = 0,531
This means that in 90 percent of all possible cases, the allele is extinguished after 15 generations. A chance loss or gain of a non-adaptive allele by a population is called a genetic drift. Such a gene could, for example, enter a population via migration of individuals of foreign populations to a population. A genetic exchange between populations is also called gene flow.
Mutations are pretty common in natural populations. It is estimated that they occur with a frequency of 10 –5 to 10 –6 per gene location and generation, though the probability of occurrence varies strongly from gene location to gene location. On an average rate of mutation, the probability of exchange of one allele by another would be extraordinarily low. A decrease of, for example, p = 0.5 to p = 0.1 ( decrease from 50 to 10 percent) would take 160,000 generations. This example shows that the development of the abundance of forms as we know it in nature cannot be caused by the accumulation of neutral mutations. The situation changes drastically, when introducing selective advantages.
An advantageous dominant allele spreads very fast, while recessive alleles have an only low ability to assert themselves even if they provide a selective advantage. The size of the population is decisive. The smaller a population, the faster the establishing of a new mutation. All new species developed from small initial populations. E. MAYR (1942) called them founder populations. Inbreeding supports the establishing of mutations, but helps also to accumulate disadvantageous mutations. As a consequence, the rate of extinction of inbreeding populations as well as of ‘founder populations’ is very high.

Ability of mutations to assert themselves (yellow, red, blue, and green) in large and small populations and with asexual and sexual propagation. The combination of mutants is indicated by mixed colours. See the following text for more details ( according to J. F. CROW and M. KIMURA, 1979).
This model is based on the assumption that alleles are adaptive in three gene locations (A, B, C) and occur simultaneously. In the case of asexual propagation, an addition of A and B occurs only, if the mutation B arises anew in a population of A individuals (H. J. MULLER, 1932).
Each natural population consists of a number of genotypes (pure lines in the sense of JOHANNSEN). This means that selection has not the same effect on all individuals. As a consequence, the structure of a population changes after selection. In a strictly formal sense, several possibilities leading to different results can be distinguished (see the following illustration).
The so-called disruptive selection has the strongest effect as the average genotype and thus the majority of a population’s individuals becomes disadvantageous. It favours extreme genotypes, and the development of a two-peaked distribution can finally and with the help of several further factors lead to the development of two new species.
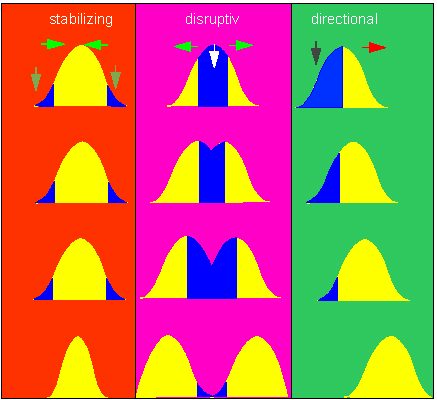
Three types of selection. 1. (red): stabilizing selection. The environmental conditions favour individuals corresponding with the average population. The variability within the population decreases over time. 2. (violet): disruptive selection. The selective pressure is directed against the average population. The extremes become favourable. The population splits finally into two partial populations. 3. (green): directed selection. The selective pressure is directed against individuals at one side of the distribution. The curve does therefore shift. This type is by far the most common in natural populations ( according to O. T. SOLBRIG, 1970, O.T. SOLBRIG, D.J.SOLBRIG, 1979).
|
|