

Den Asterales gehören neben einigen kleinen eine große Familie an. Mit 20 000 Arten sind
die ![]() Asteraceae (= Compositae, Korbblütler) die bei weitem arten- und
individuenreichste Familie der Dikotyledonen. Morphologisch sind sie relativ
einheitlich, was u.a. darin zum Ausdruck kommt, daß diese hohe Artenzahl
widerspruchslos in einer Familie untergebracht werden kann. Eine Variabilität
kommt auf der Ebene der sekundären Pflanzenstoffe zum Ausdruck. Die
vegetativen Teile sind oft mit Drüsenzellen besetzt. Die Kohlenhydrate
werden in Form von Polyfructosanen, meist als Inulin, gespeichert. Charakteristisch
ist die Bildung von Polyacetylenen (gebildet in Sekretkanalzellen), bitteren
Sesquiterpenlactonen, dem einen oder dem anderen Alkaloid (z.B. Pyrrolizidinalkaloiden
= Senecio-Alkaloiden) sowie einer Vielzahl von Monoterpenen und anderen
terpenoiden Geruchstoffen. Selten sind cyanogene Verbindungen, fast nie
sind Gerbstoffe anzutreffen. Der evolutionäre Erfolg der Asteridae
hat offensichtlich mehrere, miteinander zusammenhängende Gründe:
Asteraceae (= Compositae, Korbblütler) die bei weitem arten- und
individuenreichste Familie der Dikotyledonen. Morphologisch sind sie relativ
einheitlich, was u.a. darin zum Ausdruck kommt, daß diese hohe Artenzahl
widerspruchslos in einer Familie untergebracht werden kann. Eine Variabilität
kommt auf der Ebene der sekundären Pflanzenstoffe zum Ausdruck. Die
vegetativen Teile sind oft mit Drüsenzellen besetzt. Die Kohlenhydrate
werden in Form von Polyfructosanen, meist als Inulin, gespeichert. Charakteristisch
ist die Bildung von Polyacetylenen (gebildet in Sekretkanalzellen), bitteren
Sesquiterpenlactonen, dem einen oder dem anderen Alkaloid (z.B. Pyrrolizidinalkaloiden
= Senecio-Alkaloiden) sowie einer Vielzahl von Monoterpenen und anderen
terpenoiden Geruchstoffen. Selten sind cyanogene Verbindungen, fast nie
sind Gerbstoffe anzutreffen. Der evolutionäre Erfolg der Asteridae
hat offensichtlich mehrere, miteinander zusammenhängende Gründe:
Durch Verkürzung der Biosynthesewege sekundärer Pflanzenstoffe wurde die Möglichkeit eröffnet, zahlreiche neue Substanzen (Derivate einfacher Produkte) zu bilden. |
|
Dadurch wurde die mittlerweile erworbene Resistenz der Freßfeinde gegenüber iridoiden Substanzen umgangen. Neuartige Abwehrstoffe erlaubten es, sich gegenüber anderen Pflanzen durchzusetzen. |
|
Verkürzte Biosynthesewege sind energiesparend. Lignin wird meist nur in geringen Mengen produziert. Die Entwicklungsdauer der Pflanzen ist dadurch verkürzt. Sie sind dadurch für Biotope mit kurzer Vegetationsdauer präadaptiert. |
|
Alle bei den Asteraceae zu beobachtenden morphologischen Merkmale gelten als abgeleitet. Sie sind folglich optimiert, und alle Vorteile (Reduktion morphologischer Einheiten, Zusammenfassung der reduzierten Teile), die man jedem für sich auch anderen Taxa zuschreiben kann, kommen hier vereint zur Wirkung. Es gibt optimale Anpassungen an Bestäuber (es gibt aber auch Anemophilie), optimale Samenverbreitung (Verbreitung durch Wind und Exozoochorie, aber nur selten Endozoochorie (zu energieaufwendig !) und optimal funktionierende vegetative Organe. |


Edelweiß: Leontopodium alpinum. Nur wenige Köpfchen vorhanden (Bildmitte, silbrig glänzend). Sie sind zu einem Köpfchen zweiter Ordnung zusammengefaßt, das wiederum von filzig behaarten Honigblättern umgeben ist.
Die Asteraceae sind weltweit verbreitet, typischerweise sind es Pflanzen offener Habitate. Selten sind sie eigentlich nur in dichten Wäldern (und damit auch in den feuchten Tropen) - trotzdem: es gibt auch waldbildende Asteraceenarten - , selten sind sie auch in aquatischen Lebensräumen. Die Familie ist, wie schon angedeutet, morphologisch relativ leicht zu fassen. Problematisch ist es, die Vielzahl der Gattungen gegeneinander abzugrenzen. Verschiedene Rekombinationsmechanismen, wie Fremd- neben Selbstbefruchtung und / oder Apomixis , ein hohes Bastardierungspotential und die Tendenz, in neuen Lebensräumen Varianten auszubilden, erschweren das Bestimmen. Über das Beispiel Hieracium haben wir bereits gesprochen.
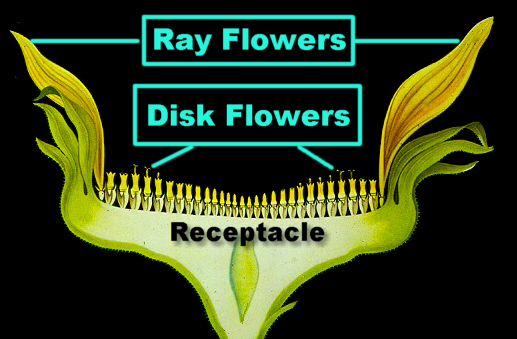
Viele Asteraceae werden als Schmuckpflanzen kultiviert, und auch die erfolgreiche Züchtung vieler neuer Sorten macht deutlich, daß das Evolutionspotential noch lange nicht erschöpft ist. Bekannte Gattungen: Taraxacum, Dahlia, Helianthus, Tagetes, Aster, Solidago, Carduus, Chrysanthemum, Cirsium u.a. Hier die wichtigsten Kennzeichen der Blütenstände: Die Blüten stehen stets zu mehreren bis vielen in von Hochblättern umgebenen, oft eine Einzelblüte vortäuschenden Köpfchen. Sie sind von einem oder von mehreren Hüllblattkreisen umgeben; die Hüllblätter sind dabei oft dachziegelartig angeordnet; bei Arten einiger Gattungen (Carlina, Cynara, Xeranthemum, Helichrysum, Gnaphalium u.a.) sind sie zum Teil gefärbt und blumenblattähnlich ausgebildet.
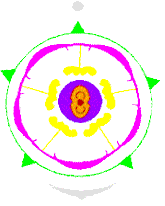 BLÜTENDIAGRAMM Arnica montana (Asteraceae) |
Die Blüten stehen auf scheibenförmig verbreiterten, kugeligen oder schüsselförmigen, meist vertieften Korbböden in der Achsel sogenannter Spreublätter, die jedoch bei manchen Arten auch fehlen. In der Regel sind die Blüten zwittrig, selten, z.B. bei Xanthium und Ambrosia, sind die Randblüten des Köpfchens eingeschlechtig. Anstelle des Kelches sind Haare (= Pappus) vorhanden, die der reifen Frucht als Flugorgan dienen. Die Blütenkrone ist entweder radiär mit fünfzipfliger, trichterförmiger Röhre (=Röhrenblüte) oder stark zygomorph (= Zungenblüte).
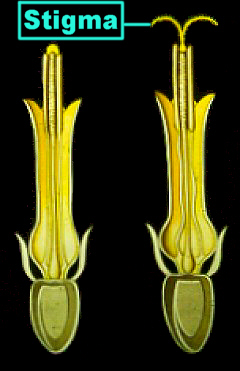
Die Staubblätter sind zu einer den Griffel umgebenden Röhre verwachsen. Die Antheren öffnen sich an ihren Innenseiten und entladen (wie bei den Campanulales) den Pollen an die mit Haaren besetzten Griffelseitenflächen. Die Blüten sind also wieder protandrisch. Der Fruchtknoten ist stets unterständig, einfächrig und aus zwei Karpellen zusammengesetzt. Die Frucht ist eine vom Pappus (wenn vorhanden) gekrönte Schließfrucht, eine Achaene, bei der Frucht- und Samenschale miteinander verwachsen sind. Die Samen sind endospermlos. Die Asteraceae können in zwei Unterfamilien untergliedert werden:
(1.) Cichorioideae (= Liguliflorae). Die Blütenkörbchen enthalten bei ihnen nur zygomorphe Zungenblüten. Darüber hinaus zeichnen sich die vegetativen Teile durch ein gegliedertes milchsaftführendes Gewebe aus, dessen Latex Triterpenverbindungen enthält. Als typisches Beispiel kann Taraxacum officinale (Löwenzahn) genannt werden.
 ( 2.) Asteroideae (= Tubuliflorae). Ihre Blütenkörbchen
enthalten entweder Zungenblüten und radiärsymmetrische Röhrenblüten
[Beispiel: Bellis perennis, (Gänseblümchen) oder nur Röhrenblüten
(Beispiel: Matricaria matricarioides, (Strahlenlose Kamille)].
Die vegetativen Organe dieser Unterfamilie sind von schizogen entstandenen,
mit Epithel ausgelegten Öl- und Harzgängen durchzogen. Lange
Zeit nahm man an, die Asteraceae (Asterales) würden sich von den Campanulales
ableiten lassen, da sie über den gleichen Mechanismus der Pollenpräsentation
verfügen, darüber hinaus Inulin als Speicherkohlenhydrat besitzen.
Aber einen solchen Mechanismus der Pollenübertragung findet man auch
bei einer Familie der Rubiales. Zudem bestehen beträchtliche Unterschiede
in der Embryonalentwicklung der beiden Ordnungen. Weil es schließlich
unter den primitiven Asteraceen auch baum- und strauchartige Arten gibt,
ist anzunehmen, daß auch die Vorfahren Holzpflanzen waren. Demnach
würden die Campanulales als bereits zu spezialisiert als Vorfahren
ausfallen. Es spricht manches dafür, daß man die Asterales in
der Nähe der Rubiales anzusiedeln hat, es wird aber, vor allem aufgrund
der starken Übereinstimmung der sekundären Pflanzenstoffe, auch
eine Verwandtschaft mit den Araliales diskutiert; doch ist auch diese nicht
gesichert.
( 2.) Asteroideae (= Tubuliflorae). Ihre Blütenkörbchen
enthalten entweder Zungenblüten und radiärsymmetrische Röhrenblüten
[Beispiel: Bellis perennis, (Gänseblümchen) oder nur Röhrenblüten
(Beispiel: Matricaria matricarioides, (Strahlenlose Kamille)].
Die vegetativen Organe dieser Unterfamilie sind von schizogen entstandenen,
mit Epithel ausgelegten Öl- und Harzgängen durchzogen. Lange
Zeit nahm man an, die Asteraceae (Asterales) würden sich von den Campanulales
ableiten lassen, da sie über den gleichen Mechanismus der Pollenpräsentation
verfügen, darüber hinaus Inulin als Speicherkohlenhydrat besitzen.
Aber einen solchen Mechanismus der Pollenübertragung findet man auch
bei einer Familie der Rubiales. Zudem bestehen beträchtliche Unterschiede
in der Embryonalentwicklung der beiden Ordnungen. Weil es schließlich
unter den primitiven Asteraceen auch baum- und strauchartige Arten gibt,
ist anzunehmen, daß auch die Vorfahren Holzpflanzen waren. Demnach
würden die Campanulales als bereits zu spezialisiert als Vorfahren
ausfallen. Es spricht manches dafür, daß man die Asterales in
der Nähe der Rubiales anzusiedeln hat, es wird aber, vor allem aufgrund
der starken Übereinstimmung der sekundären Pflanzenstoffe, auch
eine Verwandtschaft mit den Araliales diskutiert; doch ist auch diese nicht
gesichert.


© David T. WEBB, University of Hawaii at Manoa- Plant Anatomy Home Page
© Peter v. Sengbusch - b-online@botanik.uni-hamburg.de