
Wachstums- und Differenzierungsprozesse werden zusammenfassend Entwicklung genannt. Es sind Eigenarten aller vielzelligen (und einzelligen) Organismen. Bei Pflanzen setzt eine Differenzierung der Zellen unmittelbar nach der Etablierung der Polarität ein. Sie beruht auf einer ungleichen Nutzung genetischer Information in den differenzierten (spezialisierten) Zellen. Die Genaktivierung - gerade bei Pflanzen - unterliegt der Kontrolle endogener und exogener Faktoren, was bedeutet, daß Kontrollgene und Umweltfaktoren gleichermaßen die Expression zahlreicher Strukturgene beeinflussen. Selten geht es dabei um Alles-oder-Nichts-Reaktionen, meist um Steigerung oder Verminderung der Transkriptionsaktivitäten. Bei einem Wechsel in Richtung auf eine bestimmte Spezialisierung wird nicht das gesamte alte Programm stillgelegt und durch ein neues ersetzt, sondern es geht darum, das Verhältnis der gebildeten Enzyme, anderer Strukturproteine und RNS zueinander zu verändern.
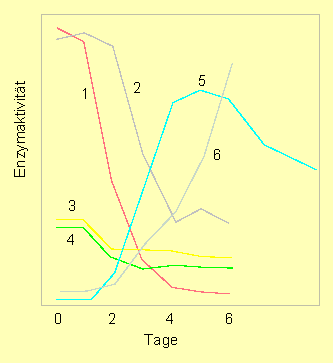
Aktivitätsänderungen von Enzymen der Purinwiederverwertung in den Koleoptilen der Lupine unmittelbar nach der Keimung. 1. Adenosinkinase, 2-4: Phosphoribosyltransferasen, 5. Adenosinnukleosidase, 6. Nukleosidphosphotransferase (C. WASTERNACK, 1982).
Wenn gesagt wird, ein oder mehrere Phytohormone steuern die Entwicklung bestimmter Prozesse, müßte man eigentlich ergänzend darauf hinweisen, daß zunächst die Enzyme produziert oder aktiviert werden müssen, die für eine Hormonsynthese benötigt werden; gleichzeitig müssen jene Gene exprimiert werden, deren Produkte für die Empfindlichkeit einer Zelle gegenüber einem Hormon zuständig sind. Nicht minder wichtig ist eine Regulation auf Translationsebene sowie auf der Ebene der fertigen oder nahezu fertigen Proteine. Die Aktivierung kann beispielsweise durch verändertes Ionenmilieu, durch Bindung von Kofaktoren, Phosphorylierung oder durch proteolytische Abspaltung eines terminalen Peptids erfolgen.
Die Aktivierung eines vorhandenen Enzyms ist an einer linearen Reaktionskinetik (als Funktion der Zeit) erkennbar. Ein Beispiel dafür ist die Aktivierung der Amylopektinase zum Zeitpunkt der Samenkeimung. Die Initiation einer de novo-Synthese erkennt man an einer sigmoiden Reaktionskinetik.
Die Verzögerung in der Anlaufphase (lag-Phase) entspricht der Zeitspanne, die für eine Enzymbildung benötigt wird. Ein Beispiel für diesen Fall wäre die alpha-Amylase, deren Synthese durch die Samenkeimung induziert wird.

Nach der Keimung (Pisum sativum) steigt die Aktivität einiger Enzyme (z. B. die der Amylopektinase) rasch an. Die Aktivitätszunahme wird nicht durch Transkriptionshemmstoffe (z. B. Chloramphenicol) beeinflußt. Folglich muß das fertige Enzym oder dessen mRNS bei der Keimung bereits vorhanden sein. Andere Enzyme, z. B. die alpha-Amylase erscheinen erst viel später. Ihr Erscheinen kann durch Chloramphenicolzugabe unterbunden werden. Ihre Synthese setzt also erst im Anschluß an die Keimung ein (Y. SCHAIN, A. M. MAYER, 1968).
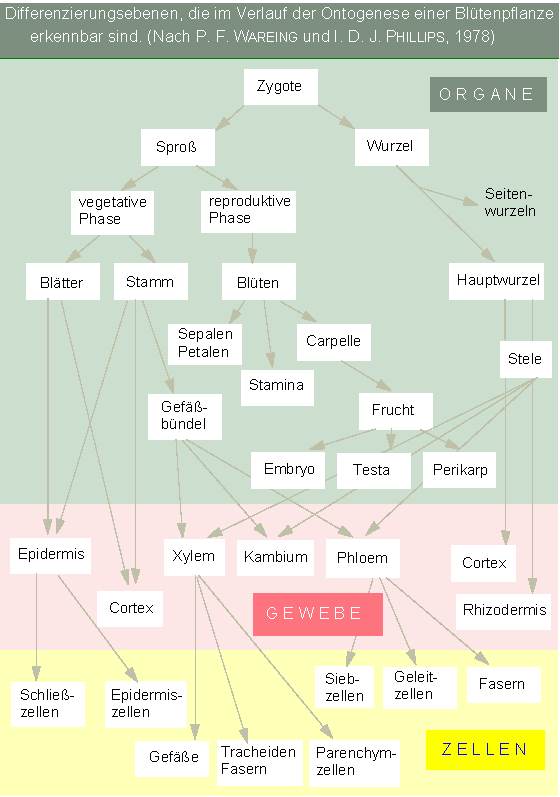
Die Entwicklung (Ontogenese) beginnt mit einer Untergliederung des Embryos in Organanlagen, aus denen Organe entstehen, in denen wiederum die Zellen (zu Geweben zusammengefaßt) sich weiter spezialisieren. Die Entwicklung einer höheren Pflanze kann daher in drei Phasen untergliedert werden:
| Juvenile Phase, beginnend mit der Samenkeimung. | |
| Wachstums- und Reifephase. Sie umfaßt das vegetative Wachstum, einschließlich der Ausbildung der Organe und Gewebe, und reicht bis zur Entfaltung der Reproduktionsorgane. | |
| Reproduktive Phase und Seneszenz. |
Die Samenkeimung kann, wie eingangs vermerkt, durch mehrere externe Signale induziert werden, wenn im Samen selbst eine Keimungsbereitschaft vorhanden ist. Die Keimung setzt daher zum günstigsten Zeitpunkt einer Vegetationsperiode ein; die Samenruhe (Dormanz) wird durchbrochen. In Regionen mit saisonbedingten Temperaturschwankungen sowie sich periodisch ändernden Tageslängen sind die Kontrollmechanismen einfach und zuverlässig (Photoperiodismus), während in Wüsten mit ihren nur spärlichen, sporadisch auftretenden Niederschlägen andere Strategien verfolgt werden müssen. Es gibt dort Arten, die die Aufeinanderfolge von Niederschlägen addieren und deren Samen erst nach Überschreiten einer bestimmten Schwelle von Regentagen keimen. Die Schwellen sind individuell unterschiedlich, so daß nicht gleich nach den ersten Regenfällen alle Samen keimen. Die früh keimenden wahren ihre Chance, indem sie jeden Regentag für ihre Entwicklung nutzen, die mit höherer Schwelle warten auf ggf. bessere Zeiten.
In ruhenden Samen erfolgt keine DNS-Replikation und die Proteinbiosynthese bewegt sich auf einem gerade noch nachweisbaren Niveau. Nach der Keimung, die fast immer mit einer durch Wasseraufnahme bedingten Volumenzunahme der Zellen verknüpft ist, setzen zahlreiche Stoffwechselaktivitäten ein. Die Nukleinsäure- und Proteinbiosynthese nehmen sprunghaft zu, der Aktivitätsspiegel der dafür benötigten Enzyme ändert sich dramatisch, die Chlorophyllsynthese setzt nach Belichtung der Sproßachsenanlage ein; die Umwandlung der Proplastiden in Plastiden erfolgt, die Photosynthese kann beginnen.
Während der Wachstumsphase erreichen die Stoffwechselleistungen
ihre Optima. Die Folge ist eine rasche Zunahme an Biomasse; Wachstum
ist daher stets mit einer irreversiblen Gewichtszunahme verbunden,
die Zellen nehmen an Zahl und Volumen zu. Der Sekundärstoffwechsel
setzt in der Regel dann ein, wenn der Primärstoffwechsel
(vor allem die Photosynthese) auf
vollen Touren läuft und genügend Assimilate zur Weiterverarbeitung
akkumuliert sind. Sämtliche regulierten Wachstums- und Stoffwechselaktivitäten
durchlaufen eine exponentielle Phase der Aktivitätszunahme,
der eine Sättigungsphase folgt, in der es kaum Zuwachs gibt,
bis das Wachstum als Funktion der Zeit asymptotisch zum Stillstand
kommt. Die Leistungsreduktion ist dann an zahlreicheichen Parametern
ablesbar. Am auffallendsten ist vielleicht die Reduktion der Blattgröße.
An jedem Sproß sind die untersten, zuerst gebildeten Blätter
größer als die an der Spitze stehenden (jüngsten).
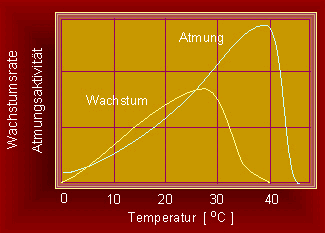
Wachstumsrate, Aktivität physiologischer Reaktionen und Temperatur sind miteinander gekoppelt.
Der Übergang zur reproduktiven Phase wird an anderer Stelle im Zusammenhang mit dem Problem Blühhormon detailliert diskutiert. Die Blütenbildung selbst kann ihrerseits in drei Abschnitte untergliedert werden:
- Induktion
- Initiation eines Blütenmeristems
- Anthese, d.h., Entfaltung der Blüte
Der zeitliche Verlauf der Bildung einzelner Blütenorgane (Staubblätter, Fruchtblätter) ist bei den einzelnen Arten (der Angiospermen) unterschiedlich geregelt, mehr dazu im nächsten Abschnitt. Bei der Mehrzahl der Arten reifen Staub- und Fruchtblätter gleichzeitig. Bei einigen Arten reifen die männlichen Blütenorgane zuerst (protandrische Blüten), bei anderun die weiblichen (protogyne Blüten). Bei einigen Arten, wie dem Kürbis (Cucurbita pepo), erscheinen zuerst männliche Blüten, dann zwittrige, schließlich weibliche, und ganz zum Schluß parthenokarpe, in denen sich auf ungeschlechtlichem Wege Samen bilden.
Mit der Blütenbildung setzt auch die seneszente Phase ein, in der die Assimilate mobilisiert und für die Samenbildung eingesetzt werden. Darüber hinaus werden sie in überdauernde Organe (sofern vorhanden) kanalisiert. Die vegetativen Organe stellen ihre Stoffwechselaktivitäten sukzessive ein und beginnen (sofern sie nicht mehrjährig sind) zu welken; absterbende Teile der Pflanze werden abgestoßen.
Die bisher beschriebenen Schritte gelten in erster Linie für die einjährigen Pflanzen, mit Einschränkungen aber auch für die mehrjährigen, beschreiben dort allerdings nicht alle zu beobachtenden Abläufe. Bei einer mehrjährigen Pflanze, z.B. einem Baum, ist zwischen den überdauernden Teilen, deren Anteil durch jährlichen Zuwachs zunimmt, und den nur eine Vegetationsperiode lebenden zu unterscheiden. Die Bildung vegetativer Sprosse und Blüten an der Peripherie des Vegetationskörpers unterliegt den gleichen Gesetzmäßigkeiten wie bei den annuellen. Anstelle der Samen wären hier die Knospen zu nennen, denn auch sie repräsentieren einen Ruhezustand, der ebenfalls durch ein Zusammenspiel von exogenen und endogenen Faktoren gebrochen werden muß.
Der Begriff Homöose wurde 1894 von BATESON geprägt, womit alternative Entwicklungswege eines bestimmten Organs beschrieben werden. Doch schon J. W. v. GOETHE stellte 1790 fest, daß Pflanzen aus einer Serie äquivalenter (homologer) Organe bestehen und daß eine progressive Transformation - er nannte sie Metamorphose - im Verlauf der Entwicklung eintritt, als dessen Folge die einzelnen Organe, beginnend mit Blättern und endend mit den Teilen der Blüte entstehen. Er nahm an, daß für den Fortgang der Entwicklung ein zunächst roher Saft verantwortlich sei, der zunehmend gereinigt würde, so daß neue Organe entstehen könnten. Als Belege für Umwandlungen führte er den Ersatz von etwa Staubblättern durch Blütenkronblätter an, so wie man das bei "gefüllten" Blüten allenthalben beobachten konnte. Gärtner gaben sich schon immer viel Mühe, solche Formen zu selektieren.
Nachdem die Methoden der Molekularbiologie etabliert waren, wandte man sich zunächst wieder der Entwicklung von Drosophila zu und stellte fest, daß es eine Klasse von Genen gibt, die darüber entscheiden, ob und wo bestimmte Kompartimente (Flügel, Augen usw.) initiiert werden und ob eine Organanlage in eine andere umgewandelt wird. Solche Gene wurden homöotische Gene genannt, und wie man heute weiß, kommen sie bei allen Pro- und Eukaryoten vor und steuern deren Entwicklung (COEN, 1991, GASSER, 1991). Frau C. VOLLHARD-NÜSSLEIN vom Tübinger Max-Planck-Institut erhielt für ihre Beitäge auf diesem Gebiet den Nobelpreis für Medizin des Jahres 1995. Homöotische Gene greifen in den Transkriptionsprozeß ein, es sind demnach Schalter, die den Satz von Genen en bloc aktivieren, der zur Ausbildung des betreffenden Kompartiments (Organs) benötigt wird.
Es lag daher nahe, auch bei Pflanzen gezielt nach solchen Genen zu suchen. dem kam zugute, daß zur Klärung molekularbiologischer und entwicklungsbiologischer Probleme ein neues Versuchsobjekt eingeführt wurde, eine kleine unscheinbare Cruciferae, Arabidopsis thaliana (MEYEROWITZ und PRUITT, 1985). Die Generationsdauer beträgt unter Laborbedingungen nur 4-5 Wochen, sie benötigt nicht viel Platz, sie besitzt eines der kleinsten pflanzlichen Genome: siehe dazu die Gendatenbank von Arabidopsis thaliana unter:
Es ist relativ einfach eine Vielzahl von Mutanten zu gewinnen, bei denen der Hormonhaushalt und einzelne Entwicklungsschritte beeinflußt sind. Die Genomgröße beträgt ca. 2 x 108 Basenpaare. Es wird zur Zeit in mehreren Arbeitsgruppen intensiv daran gearbeitet, die Nukleotidsequenz aufzuklären und man kann damit rechnen. daß die Arbeiten in Kürze abgeschlossen sein werden, so daß dann die erste vollständige genetische Information eines pflanzlichen Genoms bekannt sein wird.
Besonders aufschlußreich waren die Arbeiten aus dem Labor von E. M. MEYEROWITZ (California Institute of Technology, Pasadena) zur Determinierung der einzelnen Blütenkreise während der Anthese (Blütenbildung). Die Mutante apetala 2 bildet keine Petalen und Sepalen, die Mutante apetala 3 und einige andere bilden keine Petalen und keine Staubblätter und die Mutante agamous bildet Sepalen und Petalen, aber weder Staubblätter noch Karpelle. Diese Gene werden MADS-box Gene genannt. Zusammenfassend lassen sich die Befunde wie folgt deuten. Zur Bildung der Sepalen und Petalen wird eine Genaktivität benötigt, die A genannt wurde, zur Ausbildung von Petalen und Stamen wird eine Genaktivität B benötigt, und zur Ausbildung von Stamen und Karpellen eine Genaktivität C; das heißt, daß zur Ausbildung von Petalen und Stamen zwei Gene benötigt werden und daß der Wechsel von einem Blütenkreis zum nächsten auf dem sukzessiven Einschalten der entsprechenden Schalter beruht. Nahezu identische Ergebnisse erhielt die Arbeitsgruppe von H. SAEDLER am Max Planck Institut für Züchtungsforschung in Köln (SCHWARZ-SOMMER et al., 1990), durch Arbeiten an Anttirrhinum majus. Die genetischen Befunde wurden durch Lokalisation der entsprechenden spezifischen mRNS in den Primordien (Anlagen) für die einzelnen Blütenkreise verifiziert.

Die MADS box enthält Gene, die sich durch ein extrem konservatives Sequenzmotiv auszeichnen. Funktionell sind die Genprodukte als Transkriptionsfaktoren einzustufen. Ihre Bezeichnung MADS geht auf die Initialen der ersten vier analysierten Gene dieser Familie zurück: MCM1, AGAMOUS, DEFICIENS and SRF (serum response factor).
MADS-box Gene determinieren die Entwicklung von Infloreszenzen und Blüten, so beeinflussen sie u.a. die Entstehung der Blütenmeristeme oder jener Meristeme. die zur Differenzierung in andere Organe benötigt werden. Zunächst wurden MADS-box Gene vornehmlich an Modellbeispielen wie Antirrhinum und Arabidopsis untersucht. Die Forschung konzentriert sich inzwischen aber auch auf Fragen der Evolution und möglicher Anwendungen in der Züchtungsforschung. Es wurde dadurch möglich, ein phytlogenetisches Schema zu erstellen, das mit dem Verwandtschaftsgrad der untersuchten Arten in Einklang steht. Zu den untersuchten Monokotyledonen gehören so wichtige Kulturpflanzen wie der Mais (Zea mays ssp. mays), eine als Zierpflanze bekannte Lilie (Lilium regale) sowie die Tulpe (Tulipa gesneriana), ferner die Gymnosperme Gnetum gnemon, die Farne Ceratopteris und Ophioglossum, und das Moos Physcomitrella patens.
Weitere Einzelheiten sind auf "The MADS-box Gene Home Page" zusammengestellt, die von Günter THEISSEN's Research Group am Max-Planck-Institut für Züchtungsforschung in Köln entwickelt und betreut wird.
Homöotische Gene steuern auch die normale Entwicklung und Form eines Keimlings (Embryogenese), z.B. das apikal-basal Muster entlang der Polaritätsachse und das Radialmuster bei der Anlage der primären Gewebe sowie deren Formveränderungen (Arbeiten aus dem Labor von G. JÜRGENS, Lehrstuhl für Genetik, Universität München: MAYER, U. et al., 1991)
|
|