|
|
|
|
|
DEVELOPMENT |
|
Seeds and germination Germination in some seeds is promoted by ethylene. This could be related to an ecological response to fire which occurs naturally in many biomes. Burning vegetation generates trace amounts of ethylene and clears the ground for colonization by new plants. Many seeds require light for germination; much of our understanding of phytochrome physiology derives from experiments with lettuce seeds in which germination is stimulated by red light and inhibited by far-red. Under phytochrome control seeds that are buried in the soil will not germinate unless the soil is turned over so that they are exposed to light. This is part of the theory behind"no-till" crop production and recent ideas that it might be a good idea to plow at night! |
|
|
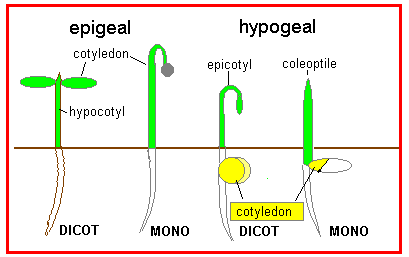
The radicle is usually the first part of the embryo to emerge from the seed coat. After this different structure emerge, depending on the species. Many (but not all) dicots are epigeal so that the hypocotyl extends, carrying the cotyledons above ground. |
|
|
In contrast, many monocots show hypogeal germination so that the cotyledon remains below ground; the coleoptile emerges and the true leaves break through this structure. However, some dicots show hypogeal germination so that the epicotyl extends, and in some monocots the mesocotyl extends carrying the cotyledon and attached endosperm upwards out of the ground. |
|
 In epigeal seedlings the apex of the emergent structure is bent over in a hook as long as it is below ground. This protects the meristem and eases its passage through the soil. Obstruction of seedling growth maintains hook closure and promotes lateral expansion to strengthen the emerging shoot. This seems to involve ethylene that is stimulated by the mechanical stress. |
|
|
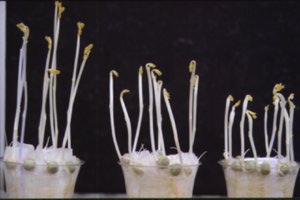
Ethylene treatment of seedlings promotes hook closure and stem thickening rather than elongation. These etiolated pea seedlings were treated with 0, 0.1 and 1 ppm ethylene (left to right) |
|
|
All the time that the seedling is in the dark it remains pale or nearly white and its leaves do not expand. When it reaches the light stem elongation is suppressed, chlorophyll synthesis and leaf expansion are stimulated under the control of the phytochrome system. When plants are covered over they return to this etiolated morphology. Chloroplasts revert to etioplasts and leaves do not expand |
|
|
|
|
|
In all plants the first stages of growth involve extension from single stem and root apices. In many dicots the radicle persists to form a tap root from which lateral roots arise. In monocots the first root is soon replaced by adventitious roots from the base of the stem. The original stem apex tends to be dominant, at least for a time. The apical buds of extending shoots suppress the growth of lateral, axillary buds. This seems to be because auxin produced in the apex travels down the stem and prevents bud-break. In contrast, auxin in the root inhibits elongation, but promotes the initiation of laterals. Roots are a sources of cytokinins that move up the plant and tend to promote shoot initiation. This is part of the system through which plants control the balance between shoot and root. Removal of the apical bud allows the laterals to grow out; this may be desirable in order to get a bushy plant or many flowers on one stem. At other times excessive bud break after pruning may be undesirable, in which case auxin paints can be used on the cut surface to maintain the suppression of laterals. |
|
|
|
|
|
The mature plant Light has a modulating effect on plant growth. Shade tends to promote elongated growth and the development of broad, thin leaves. Light filtering through foliage is enriched in far-red so that inhibition of stem elongation by Pfr is negated. The red/far-red system influences the amount of growth of stems and leaves, whereas the blue-light receptor influences direction. Thus plants in a dense stand or branches in a tree adjust their growth to position the foliage for optimum light capture |
|
|
|
|
|
Copyright © Michael Knee, The Ohio State
University |
|