
It has been mentioned in the historical outline that photosynthesis is dependent on light. The results of ENGELMANN and SACHS showed that it is absorbed by chlorophyll. We also got to know that plants have two types of chlorophyll, chlorophyll a and b and that both types display a characteristic absorption spectrum.
The action spectrum of photosynthesis resembles the absorption spectra of chlorophyll though it is not identical. This means that further photoreceptors (so-called accessory pigments) exist.
We already got to know something about the assimilation of carbon dioxide in the previous section. Our question is now: which reactions are induced by light and how is the light energy converted into chemical energy? Or, in other words, how are ATP and NADPH2 produced?
CALVIN and his collaborators studied the dark reactions in intact, active cells. This attempt proved to be insufficient for the light reactions. The results were contradictory. Techniques to isolate active chloroplasts had to be developed.
After the use of fractions containing isolated chloroplasts became usual, were three research groups at the same time (1951) and independent of each other able to show that isolated chloroplasts reduce NADP to NADPH2 when exposed to light [W. VISHNIAC and S. OCHOA (Rockefeller Institute, New York), L. J. TOLMACH (University of Chicago) and D. I. ARNON (University of California, Berkeley)]. Shortly afterwards (in 1954) discovered ARNON and his collaborators that the production of ATP, too, is dependent on light and that both ATP and NADPH2 can be produced simultaneously. Both compounds are generated from precursors that are present in the chloroplast already before photosynthesis starts, since no extern metabolites were supplied during the experiments. Accordingly was light (photons) the only available source of energy. It turned out that the production of ATP needs no oxygen, neither is oxygen produced during the reaction. Consequently runs the equation as follows:
n ADP + n Pi > (photons) > n ATP
The process is termed photophosphorylation. It exists in bacteria and blue-green algae, too, and is a general feature of photosynthetic processes.
After it was proven hat ATP is produced, was it asked how this is done. It seemed unlikely that the light induces the production of ATP directly but it had turned out that the production of ATP has to be preceded by an exposure to light. The concept of a light-induced electron flow was developed. It assumes that one molecule of chlorophyll absorbs one photon. As a consequence is an electrons of chlorophyll transferred to a higher energy level.
This energy-rich electron is then transferred to a neighbouring electron acceptor with a strong electronegative redox potential. The transfer of the electron from the activated chlorophyll to the (first) acceptor is the first photochemical phase of photosynthesis. Its decisive feature is the transformation of a photon flow (light) into a flow of electrons.
As soon as a strongly electronegative (reducing) substance has been produced can the electron flow proceed with electron acceptors of less negative redox potentials. The process releases chemical energy that is used for photophosphorylation. Already during the fifties existed the first strong proves for the involvement of the chloroplasts' cytochromes. It could be shown, too, that the electron is finally accepted by a chlorophyll, so that its original state is restored again. The requirements of catalysis are fulfilled. The process became known as cyclic phosphorylation (D. I. ARNON, 1959).
Such a cyclic flow of electrons that is powered by light and releases chemical energy used for the production of ATP is unique. It is the outstanding property of photosynthetic cells.
Concept of cyclic photophosphorylation (to the left). To the right: the original concept of non-cyclic photophosphorylation (according to D. I. ARNON, 1971)
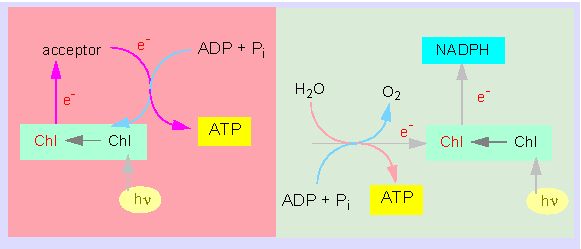
The only unexplained process remained was the photoreduction of NADP in chloroplasts. It were again ARNON and his collaborators that were in 1957 able to discover a second part of photophosphorylation. They could prove experimentally that the photoreduction of NADP and the synthesis of ATP are coupled. In contrast to the cyclic photophosphorylation is the production of ATP coupled stoichiometric to a light-induced transfer of electrons from water to NADP and to the production of oxygen. The ATP production of the whole system increases the reduction rate of NADP. This pointed at the fact that the process is tightly coupled to cyclic photophosphorylation. Since electrons are irreversibly transferred from chlorophyll to NADP, are substitutes needed and these electrons stem from the breakdown of water. It is spoken of non-cyclic photophosphorylation, since ATP is produced simultaneously. Ferredoxin (a heme-less iron-sulphur protein) has a key position in this process. Its reduction potential is far more negative than that of NADP so that an electron flow from ferredoxin to NADP was very likely. The reduction of NADP is a three step reaction:
- a photochemical reduction of ferredoxin that is followed by two 'dark' steps.
- The re-oxidation of ferredoxin with the help of a ferredoxin-NADP-reductase (a flavoprotein).
- The re-oxidation of the ferredoxin-NADP-reductase by NADP.
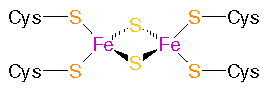
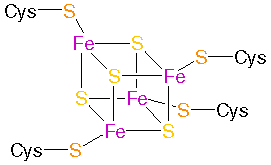
Ferredoxin: Iron-sulphur-complex: - to the left: Fe2S2 protein - Plant�type ferredoxins , to the right:
Fe4S4 proteins - Bacterial�type ferredoxins
from: PROMISE - The Prosthetic groups and Metal Ions in Protein Active Sites Database
What was at first regarded as a photoreduction of NADP proved to be an electron transport chain that runs from ferredoxin via a flavin component to NADP.
The outstanding position of ferredoxin was strengthened even more after it was found out that stoichiometric amounts of O2 and ATP are produced during the reaction. The non-cyclic photophosphorylation can accordingly be described as follows:
4 ferredoxin (oxidized) + 2 ADP + 2 Pi + 2 H2O > (photons) > 4 ferredoxin (reduced) + 2 ATP + O2 + 4 H+.
The results led to the question how this reaction is coupled to the cyclic photophosphorylation discussed at the beginning. A series of experiments using specific inhibitors showed that ferredoxin is a component of that pathway, too.
So much about the chemical data. More knowledge about the primary effect of the light and about the significance of chlorophyll would have to exist to interpret them. These problems, too, have a long past history.
In 1932 exposed R. EMERSON and U. ARNOLD of the University of Illinois at Urbana Chlorella cells to a series of extremely short flashes of light. With this experiment did they try to find out how many molecules of chlorophyll were necessary to use one photon for the production of one molecule of oxygen. The result was that several hundred chlorophyll molecules are necessary which means that not all of them are of the same importance. Most act as light traps (or antennas) helping to transfer a photon to a reaction centre where an especially exposed chlorophyll transforms light energy into chemical energy. H. GAFFRON called this complex of several hundred chlorophyll molecules and other pigments (carotenes, carotenoids, xanthophylls, etc.) a photosynthetic unit.
This aggregation of pigments seems to lead to an especially efficient use of the incoming light. Still, a rather large part of the irradiated energy is lost. It does never reach the reaction centre and is emitted as warmth or light (red autofluorescence of chlorophyll).
When regarding the absorption spectrum of photosynthesis does it stand out that the efficiency of light of the wave length lambda > 680 nm decreases strongly although chlorophyll a displays an absorption in that area. R. EMERSON (1957) discovered that light of the wave length lambda > 700 (710) increases the rate of photosynthesis drastically if light of the wave length lambda = 680 nm (or less) is present at the same time.
When these two light qualities are used independent of each other or one after the other is no increase measured. EMERSON concluded that two photochemical processes have to exist that consist of different pigment systems (light receptors) but that do co-operate (EMERSON-effect). According to a suggestion of L. N.M. DUYSENS are the two systems called
| photosystem I (PS I). It needs light of longer wave lengths (lambda > 700 nm) and | |
| photosystem II (PS II). It becomes active when exposed to shorter wave lengths (lambda < 680 nm) |
The ratio of chlorophyll a to chlorophyll b is higher in PS I than in PS II. The question how the two systems co-operate and how they are coupled to the production of ATP and NADPH2 remains to be settled.
ARNON and his collaborators could prove that the two systems are arranged in series and that both systems are required to explain all effects that had been recognized as photosynthetic ones. Only some bacteria that produce no oxygen during photosynthesis lack photosystem II. This results hints at the suggestion that the splitting of water is coupled to photosystem II and that photosystem I developed earlier in evolution.
The reaction centre of every photosystem is represented by one molecule of chlorophyll a each (P 700 in PS I and P 680 in PS II, where P means pigment).
The absorption of a photon by P 680 (which has a positive redox potential of + 0,8 V in its basic state) transfers P 680 into its excited state ( with a redox potential of 0,0 V) and causes the formation of a strongly oxidizing component (Z+) and a weakly reducing (Q-) one. Z+ withdraws electrons from water so that O2 and protons are set free.
4 Z+ + 2 H2O > 4 Z + 4 H+ + O2
The reducing component (a membrane-bound plastoquinone) feeds the electron into an electron transport chain in the course of which it looses energy part of which is used for the production of ATP. The electron does not return to its starting point (chlorophyll P 680) but is transferred to a chlorophyll molecule of photosystem I (P 700). The two photosystems are thus coupled.
The absorption of a further photon excites the P 700 just mentioned (the redox potential of which is + 0,4 - + 0,5 in its basic state). It transfers one electron to a membrane bound ferredoxin (P430) which passes the electron on to a soluble ferredoxin. The following steps are known.
In a stoichiometric sense is the outline above incomplete since ferredoxin transfers just one electron at any given time while two electrons are needed for the production of one NADPH2. The equation would consequently have to be:
2 ferredoxin (reduced) + 2 H+ + NADP+ > 2 ferredoxin (oxidized) + NADPH2
In summary is the light energy used for the flow of electrons from water to NADPH2 and for the simultaneous production of ATP (Z-scheme).
During our discussion did we neglect the cyclic photophosphorylation mentioned at the beginning. It proved to be a parallel process that starts work as soon as enough NADPH2 but too small amounts of ATP are present. Only photosystem I participates in cyclic photophosphorylation.
© Peter v. Sengbusch - b-online@botanik.uni-hamburg.de