
C. DARWIN worked for quite some time on botanical problems. His
research was on self- and foreign pollination, on the interbreeding
of different species and the registration of the number of their
offspring, though he did not draw conclusions. Besides others, he
worked with snapdragon (Antirrhinum majus), a species of the
Scrophulariaceae family. The flowers of most species of this family
are zygomorphous, i.e.
characterized by just one plane of symmetry. Only few species are of
radial symmetry as most angiosperm flowers are. But sometimes radial
symmetry does also occur in the flowers of snapdragons and other
species, e. g. Digitalis purpuraea. After
isolation and self-pollination (with pollen of the same individual),
a homozygous line can be established. DARWIN crossed such plants with
zygomorphously flowering partners and got the following result:
Zygomorphy and Radial Symmetry in Snapdragons (Antirrhinum majus) |
|||
|
parental generation |
|
||
|
|
|
||
|
|
zygomorphous |
intermediate |
radial symmetry |
|
|
|
|
|
If the intermediate and the zygomorphous individuals are counted,
then the resulting ratio is 3:1. Zygomorphy is obviously a dominant
character, but the dominance seems to be imperfect, because a weak
tendency to intermediate expression is observable.
C. CORRENS analyzed a huge number
of species and features. In a study published in 1902, he describes a
segregation of zea mays:
round seeds (dominant) : wrinkled seeds (recessive) results in a F2 generation of 8975 : 1711.
A 3:1 segregation would have resulted in a ratio of 8015 : 1711.
The deviation seemed to be too large and CORRENS explained it by a
more or less easy generation of one or the other germ cell
combination. This statement is insofar important as the Mendelian
laws base on statistic assumptions and presume that all types of
gametes occur in always the same numbers and all combinations with
the same probability. But living systems cannot always be regarded as
equal as the following examples will illustrate. DARWIN's selection
hypothesis of evolution for example, is, as you may know, founded
on the existence of unequal genotypes in one population.
A further specimen that CORRENS worked with, was the stinging nettle. The species Urtica pilulifera has leaves with serrated, Urtica dodartii such with smooth margins. Serrated is dominant. A clear 3:1 ratio does therefore appear in the F2 generation . This example has been cited in numerous textbooks and the scheme has been depicted just as often. The heterozygous plants with serrated leaves do obviously look the same as the homozygous ones. Nevertheless do the first two leaves (and only these) of heterozygous plants display a reduced number of dents that distinguishes them from the respective leaves of homozygous forms. CORRENS called this a hidden tendency towards an intermediate state.
In the classic Mendelian example, the garden pea with round or edgy seeds, a clear dominance of one seed shape can be observed. The same gene does also influence the form of the starch grains. In round seeds can simple, large longitudinal grains be found, those of edgy seeds are strangely shaped. Both forms occur simultaneously in heterozygous individuals of both the F1 and the F2 generation (A. DARBISHIRE, 1911). They do thus represent an intermediate type. The phenomenon that one gene has an influence on several characters is called pleiotropy.
It is often underestimated how large the scientific knowledge of
those that shaped and influenced genetics right from its beginning
were. Before and after the turn of the century, a whole range of
hybridizations was performed. However, only few examples did find
their way into the textbooks and it may therefore be that the wrong
impression that only these experiments were performed and analyzed
may arise. Everything that could be shown in plant experiments, could
also be verified for animals. The ever same examples depict the
conditions best and demonstrate, partly at least, seeming
deviations.
In addition, the term 'character' or 'feature' is not clearly
defined. It is solely dependent on the observer, what is meant by it
and not always is the alternative as lucid as in red and white
flowers. It is well-known that there exist numerous shades and
transitions with many flower colours and for all these phenomenons,
explanations had to be found. Farmers and plant breeders are
particularly interested in the 'yield', a character that, as every
practitioner knows, is subject to a very high degree of
variability.
The definition of 'character' is not restricted to phenotypes that can be perceived without microscopic aid. The microscope as well as other physical and (bio-)chemical techniques of analysis became an as important device for geneticists as the mathematical analysis of the results. The more it was worked on the question, what a gene is and how it exerts its effects, the more had the methods to be improved, the more important did it become to analyze an object as simple as possible and the more previous knowledge was necessary to understand the complex process step by step.
Heterostyly, again, is a phenomenon that was among those examined
by DARWIN. Many primulas are characterized by heterostyly.
About half of the flowers displays a long, the other half a short
style. The anthers are usually never at the same level as the style.
Primulas are without exception cross-fertilized and normally are
individuals with long styles only pollinated by short-style ones and
vice versa, whereby an effective protection against self-pollination
is given. In a few species (like Primula sinensis),
self-pollination can be triggered, if no foreign pollen is present.
Individuals with long styles produce only offspring with long styles
upon self-pollination, while self-pollinating primulas with short
styles produce offspring with short styles and such with long styles
in the ratio 3:1. The crossing of long styles with short styles (as
it is natural) results in both forms in the ratio 1:1.
This result can simply be explained by the fact that short styles
are caused by a heterozygous state, long styles by a homozygous one.
The keeping of the 1:1 ratio is caused by the permanent reciprocal
crossings.
This experiment points out that there exist by no means only homozygous forms in nature and that many phenomenons can be traced back to heterozygous conditions. The dominant homozygous forms of many species do hardly ever appear under natural conditions.
The picture shows that the difference of both types of flower is not only in short or long styles, but that also the surface of the style and the shape and size of the pollen are different. This means that we do additionally have to deal with an example for pleiotropy, with a number of coupled characters or with both.
The high variability of its flower colours has made the snapdragon a popular ornamental plant and gardeners have always had an interest to gain especially attractive individuals as homozygous lines. First attempts go back to the 17th century, TOURNEFORT discussed it around 1700 and in the 19th century, numerous varieties were widespread. The import and use of new varieties increased the amount of different forms. More about Antirrhinum:
Kaiser-Wilhelm-Institut für Züchtungsforschung founded in 1927 (today's Max-Planck-Institut für Züchtungsforschung in Köln) chose the primula as one of his main specimens for the analysis of the variability of flower colours. But we will start with another of his experiments.
There exists a snapdragon variety with golden ('aurea') instead of normal green leaves. If such plants are crossed, then green and golden 'aurea'-phenotypes in the ratio 1:2 result. In addition, ivory seedlings can be observed that die shortly after germination and do therefore have no influence on the mentioned ratio. E. BAUR concluded that the 'aurea' phenotypes are heterozygous, the green ones are homozygous and that the ivory seedlings represent the homozygous recessive form. The latter are not capable of surviving (the gg combination of alleles is lethal) and do therefore not appear in a grown-up state. In this case, the lethality is easy to explain. The pale appearance is obviously caused by the absence of chlorophyll and as soon as the reservoir of nutriments is used up, the development is stopped, because the plants cannot feed any more. Generally, lethality can be traced back to unfavourable combinations of genes or alleles (lethality factors) that get a look-in at different states of development.
The American geneticist A. J. MANGELSDORF identified 19 different lethality factors of corn (Zea mays) up till 1926. A range of such varieties is a suitable basic material for studies on developmental physiology.

Effect Times of a Number of Lethal Factors in Corn (yellow bars; the marking would be de1, de2, de3,........developmental mutant) (according to A. J. MANGELSDORF, 1926)
A close analysis with modern techniques allowed conclusions concerning the time point and way, in which single genes influence the development. Lethality is a relative term. It means that an individual cannot survive under certain environmental conditions, though it may develop normally under altered conditions. Plants without chloroplasts, for example, can survive, if cultured on sugar-containing substrates and with addition of growth factors.
We did already get to know an example for pleiotropy, where a gene influences several features. The Swedish geneticist H. NILSSON-EHLE wrote in 1909 about a cross of a dark-seeded and a light-seeded oat variety. He found only dark seeds in the F1 generation, in the F2 generation he obtained 630 dark and 40 light seeds. This represents a ratio of 15:1. He did now assume that two independent genes (N, M) caused the dark colour either each for itself or the two in combination with each other. In the F2 generation do thereby develop 15 individuals in a dihybrid scheme, in which at least one of the genes is present in a heterozygously dominant condition. All genotypes that contain at least a dominant N or M lead to the development of dark seeds, only a doubly recessive homozygous condition causes light seeds. Both genes have thus effects of the same kind.
The American geneticist G. H. SHULL got a comparable result, when he crossed two shepherd's purse-species, Capsella bursa-pastoris and Capsella heegeri. The first is characterized by heart-shaped little pods, the latter by oval ones. In the F1, all pods are heart-shaped, in the F2, one oval pod per 15 heart-shaped ones appears. Both genes have the same influence on the fruit shape, one can substitute for the other.
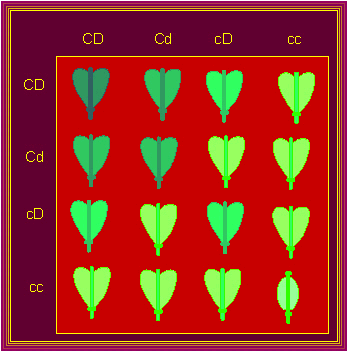
The Inheritance of Fruit Shapes in Capsella: PUNNETT-Square. The abscissa displays the genotype of the haploid gametes of one, the ordinate those of the other parent: CD, Cd, cD, cc. This results in the combinations (first row of the square): CDCD, CDCd, CDcD, CDcc. The combinations of the second row would accordingly be CdCD, CdCd, CdcD, Cdcc, etc. The colours of the diagram represent the different phenotypes (G. H. SHULL, 1914).
12 + 3 + 1 is 16. The cross of beans with dark seeds and such with white seeds results in a uniform dark F1 generation that segregates in the F2 generation not only into dark and white seeds, but also into brown ones. The ratio is 12 dark ones : 3 brown ones : 1 white bean. It is obvious that this is again a variation of a heterozygous hybrid, with the difference that the respective genes are not of the same kind. A biochemical characterization of the colour composition showed that the dark staining is caused by two different substances, both of which are dark. The dominant allele of the gene A causes the formation of a dark dye, the respective allele of B the brown one. A is present 12 times. This is also due for B, but the presence of the brown dye is concealed by the dark colour of A (in 9 of 12 individuals).
It is generally spoken of epistasy, if a gene conceals the effects of another. This term is only valid under certain conditions. If the colour composition of the single seeds is analyzed with the aid of biochemical (chromatographic) methods, with which the two dyes can be separated and analyzed independently, then an independent distribution of 3:1 can be found for each of them. Epistasy may not always be that simple. It can also cause the total suppression of a further gene (a hypostatic gene).
The last examples (15:1, 12:3:1) could be explained by the segregation of independent genes. A ratio of 9:7 in F2 is found in dihybrid crossings, where two genes that are dependent on each other have a complementary effect, i.e. where one cannot display its effect without the other. KAPPERT found that certain pea species with violet pods result in a 9 (violet) : 7 (green) segregation in the F2, if crossed with green-podded peas.
It is thus not enough, if just one gene is present in a dominant state. A biochemical analysis of such phenotypes that was performed later, offered a plausible explanation. The formation of the violet colour occurs in two steps. In the first step (caused by the gene V), a colourless product is assembled from a colourless precursor. Gene U enables its transformation into a violet dye. If V is double recessive, U cannot exert its effect, because the necessary product is missing. And if U is double recessive, the colourless precursor is produced, but it cannot be transformed into the violet dye.
|
|