
Wie im letzten Abschnitt dargelegt, ist ein Energiefluß ein wesentliches Merkmal eines jeden Ökosystems, denn alle lebenden Systeme sind als offen zu beschreiben. Sie sind auf eine ständige Energiezufuhr angewiesen, um die strukturelle Organisation und die lebenserhaltenden Funktionen aufrechtzuerhalten, denn nach dem zweiten Hauptsatz der Thermodynamik strebt jedes System den Zustand höchster Entropie an. Die Umkehr - also die Bildung entropiearmer (=geordneter) Einheiten - erfordert die Zufuhr von Energie aus der Umgebung, und da nur Pflanzen (und einige Prokaryoten, vornehmlich die Cyanophyta) Lichtenergie verwerten können, fällt ihnen in jedem natürlichen Ökosystem eine Schlüsselrolle zu.
Weil der Energiefluß ein gerichteter (vektorieller) Prozeß ist und weil beim Übergang von einer trophischen Ebene (Stufe) zur nächsthöheren bestenfalls 10 Prozent der Biomasse verwertet werden kann, nimmt die Biomasse von Trophieebene zu Trophieebene drastisch ab. In der Natur kann es daher selten mehr als vier, höchstens fünf Trophieebenen geben. Die Biomasse aller Carnivoren zusammengenommen, ist stets geringer als die der Herbivoren, und die wiederum ist geringer als die der Pflanzen. Wir erhAlten dadurch eine Nahrungspyramide, deren Gestalt von der Produktivität und Artzusammensetzung des jeweiligen Ökosystems abhängt. Die lineare Abfolge der einzelnen Glieder (Produzenten, Konsumenten 1. Ordnung ...) nennt man Nahrungskette, doch weil die Verhältnisse in natürlichen Ökosystemen meist komplexer sind, ist es besser, von einem Nahrungsnetz (oder Nahrungsgefüge) zu sprechen. Nahrungspyramiden beziehen sich in der Regel auf Angaben der Biomasse, nicht so sehr auf Arten- und Individuenzahlen.

Über 90 Prozent der Gesamtbiomasse der Erde entfällt auf Pflanzen, nur wenige Prozent auf die übrigen Organismengruppen. Der Vergleich der Artenzahlen hingegen ergibt, daß es etwa zehnmal so viele Tier- wie Pflanzenarten gibt.
Unter gewis3en Bedingungen können umgekehrte Nahrungspyramiden beobachtet werden. Die Ursache hierfür liegt entweder in einer zeitlichen oder räumlichen Versetzung des Erscheinens der einzelnen Systemelemente.
Eine Anzahl von Tieren ernährt sich von toten Pflanzen oder
Pflanzenresten. Die Biomasse lebender Tiere kann daher in manchen
Ökosystemen im Winter höher als die der lebenden Pflanzen
sein. Während der übrigen Jahreszeiten gelten die üblichen
Bedingungen. Eine räumliche Trennung ist für die Tiefsee
typisch, denn Tiefseetiere leben in Zonen, in denen Photosynthese,
und damit das Vorkommen von Pflanzen ausgeschlossen ist. Sie ernähren
sich von einem ständigen Regen abgestorbener Pflanzen (fast
ausnahmslos einzellige Algen). Um die Größenordnungen
der Energieflüsse in Ökosystemen zu ermitteln, muß
man sich zunächst mit dem Energieinput d.h., der Menge und
Qualität des Sonnenlichts auseinandersetzen.
Emissionsspektrum der Sonne sowie Filterwirkungen in der Biosphäre. Weiße Kurve: Sonnenlicht., gelbe: Sonnenlicht auf Meereshöhe, orange: durch Wolken gefiltert, grün: durch Vegetation transmittiert. Die Abszisse umfaßt den Wellenbereich von 0,1 - 10 µm (logarithmische Skala) Auf der Ordinate ist die Lichtintensität wiedergegeben. (Nach D. M.GATES, 1965).
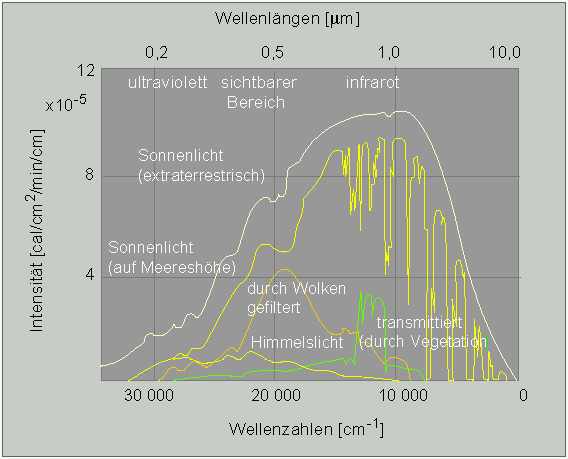
Dadurch wird deutlich, daß im Verlauf der pflanzlichen Evolution solche Pigmente selektiert wurden, die die verfügbare Lichtqualität (d.h. Wellenlänge der Strahlung) optimal nutzten. Eine Bewölkung wirkt als Wärmeschutzfilter, sie absorbiert UV- und Infrarotstrahlung; beide sind für die Photosynthese wertlos, ja sogar schädlich. Licht des sichtbaren Bereichs wird in weit geringerem Maße absorbiert. Eine Vegetationsdecke filtert sichtbares Licht. Da Chlorophylle und Phytochrom annähernd die gleichen Absorptionscharakteristika haben, gelangt hellrotes Licht kaum in tiefere Schichten. Da aber gerade dieser Wellenlängenbereich zur Steuerung von Keimungs- und Wachstumsprozessen benötigt wird, inhibiert eine dichte Pflanzendecke die Entwicklung nachwachsender Pflanzen.
Über die Erdoberfläche gemittelt (ausgenommen Polar- und Wüstenregionen) wird die Sonnenenergie mit ca. 2 cal / Quadratzentimeter / min angegeben. Dieser Wert ist die Solarkonstante. Jahreszeitliche Schwankungen sowie Unterschiede in der Exposition (Nordhang, Südhang) führen zu Differenzen im Bereich einer Größenordnung. Pro Tag erreicht demnach die Strahlungsenergie einen Durchschnittswert von 3000-4000 kcal / Quadratmeter, das sind im Jahr 1,1-1,5 x 10 000 kcal / Quadratmeter. Ein Teil dieser Energie wird von der Erdoberfläche reflektiert, ist damit für Biosyntheseprozesse von vornherein verloren. Was übrigbleibt, nennt man Nettostrahlung, für die man in Breiten zwischen 40 Grad Nord und 40 Grad Süd über dem Meer einen Wert von 1 Million kcal / Quadratmeter / Jahr und über dem Festland einen Wert von 0,6 Millionen kcal / Quadratmeter / Jahr ermittelt hat.
Wasserverdunstung und Luftbewegungen sind wichtige Faktoren, die dafür sorgen, daß der größte Teil dieser an und für sich gewaltigen Energiemengen zeitversetzt - als Wärmeenergie - ins Weltall abgeführt wird. Ohne diese Energieabgabe würde sich die Erde in kürzester Zeit überhitzen, und es würden dadurch Temperaturen entstehen, unter denen Leben nicht möglich wäre. Andererseits sind Sonneneinstrahlung und die damit verbundene Temperaturerhöhung die Hauptursache für das Auftreten von Klimazonen (tropisch, gemäßigt, polar) sowie von jahres- und von tageszeitlichen Schwankungen.
Die jährliche Biomasseproduktion wird auf etwa 164 Milliarden Tonnen geschätzt (R. H. WHITTAKER und G. E. LIKENS, Cornell University, 1975), ein Drittel davon entsteht in Ozeanen, zwei Drittel in terrestrischen Ökosystemen.
Zwischen Biomasse und Produktionsrate ist klar zu unterscheiden. Unter der Produktionsrate oder Produktivität versteht man eine pro Zeiteinheit fixierte Energiemenge. Aus vorhandener Biomasse allein lassen sich nur unter Vorgabe bestimmter Bedingungen Näherungswerte des Energieumsatzes ermitteln. Wie bereits betont, geht bei jedem Schritt im Stoffwechsel Energie in Form von Wärme verloren. Um dem Rechnung zu tragen, unterscheidet man zwischen Brutto- und Nettoproduktion. Die Nettoproduktion ist dabei der nach Abzug der durch Atmungsprozesse verlorenen Energiemenge verbleibende Restbetrag.
Zu den Schwierigkeiten der Produktivitätsbestimmung gehört das Faktum, daß oft nicht zu entscheiden ist, ob sich ein System (hier die Pflanzen) in einem Fließgleichgewicht oder in einem dynamischen Gleichgewicht (Wachstumsphase) befindet. Mit der kontinuierlichen Abnahme der Wachstumsaktivität nimmt auch die Atmungsaktivität ab, denn zum Erhalt von Strukturen wird weniger Energie benötigt als zum Aufbau neuer.
Aus der Bestimmung der Photosyntheseaktivität - gemessen an der Menge freigesetzten Sauerstoffs - läßt sich ebenfalls nur wenig über die Produktivität aussagen, denn ein Teil des Sauerstoffs wird durch die Atmung verbraucht, ein weiterer durch die Lichtatmung (Photorespiration), deren Aktivität wiederum direkt mit der zur Verfügung stehenden Lichtmenge korreliert ist. Unter kontrollierten Laborbedingungen und großem experimentellem Aufwand sind die einzelnen Faktoren voneinander getrennt meßbar. Die unter derartigen Bedingungen gewonnenen Daten können zu Proportionalitätsfaktoren umgerechnet werden. Oft bedient man sich zur Bestimmung der Biomasse der Messung des Kohlenstoffanteils (einer bestimmten Einheit). Als Umrechnungsfaktoren gelten:
10 kcal ~ 2 g Trockensubstanz ~ 1 g Kohlenstoff
Ein Energiefluß wird üblicherweise in Dimensionen von kcal / g Trockengewicht angegeben. Natürliche Ökosysteme sind auf eine maximale Bruttoproduktionsrate (hohen Umsatz) optimiert, vom Menschen beeinflußte (landwirtschaftlich genutzte Flächen) auf eine möglichst hohe Nettoproduktionsrate
Klimaxgesellschaften, z.B. der tropische Regenwald, zeichnen sich durch eine hohe Produktivität bei nahezu gleichbleibender Biomasse aus. Anders ein Ökosystem, das sich in einer Sukzession befindet. Ein Moor beispielsweise wächst durch stetige Ablagerung von abgestorbenem Sphagnum-Gewebe.
Nur etwa 5 Prozent der verfügbaren Sonnenenergie wird in Form chemischer Energie in der Biomasse der Pflanzen konserviert, davon können - rein theoretisch - allenfalls 80 Prozent von den Organismen der nächsthöheren Trophieebene genutzt werden. Die Realität sieht aber meist viel ungünstiger aus. So liegt beispielsweise in Wäldern der überwiegende Teil der Biomasse in Form von Holz fest, und das ist für tierische Ernährung denkbar ungünstig. Nur wenige Spezialisten können damit etwas anfangen. Die Beziehungen zwischen Brutto- und Nettoproduktion, sowie der Energiefluß durch Ökosysteme läßt sich am Modellsystem Wald gut demonstrieren.
Die verfügbare Lichtmenge ist in der Regel kein limitierender Faktor. Anders verhält es sich mit dem Wasser. Man denke dabei nur an das karge Pflanzenwachstum in Wüsten und vergleiche es mit der üppigen Vegetation der Tropen. Ergänzt sei, daß die Lichtmenge in den meist wolkenlosen Wüstengebieten die der wolkenbedeckten Tropen deutlich übersteigt. In bewässerten Wüstenregionen ist wegen der hohen Lichtintensität eine höhere Bruttoproduktionsrate als in Gebieten mit geringerer Lichtintensität zu erzielen. Doch aufgrund der hohen Atmungsverluste während der warmen Nächte wird von der Pflanze mehr Energie verbraucht als in kühleren Gegenden. In der Bilanz ergibt sich daraus eine Verminderung der Nettoproduktionsrate. Das erklärt, weshalb zum Beispiel Reisernten in äquatorialen Gebieten stets geringere Flächenerträge ergeben als in gemäßigten Zonen.
Die Nettoprimärproduktion kann in den einzelnen Einheiten beträchtlich schwanken, so wird der niedrigste Wert für "offenes Weltmeer" mit zwei, der höchste mit 400 angegeben, für "Kulturland" lauten die entsprechenden Angaben 100, respektive 4000 g / m2 / Jahr. In den meisten der übrigen Vegetationseinheiten schwanken die Werte um etwa das Fünffache. Zur Umrechnung von Nettoprimärproduktion auf Energiefixierung wird ein Heizwert (kcal / g) angenommen. Er liegt bei ca 4,5. Der niedrigste Wert (4,0) wird für Grasland angegeben, der höchste (4,9) für offenes Weltmeer und Auftriebszonen.
© Peter v. Sengbusch - b-online@botanik.uni-hamburg.de
