
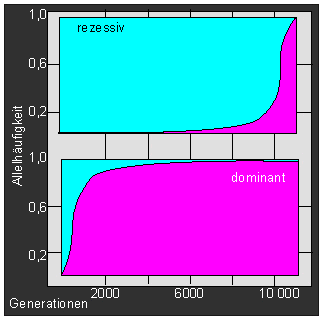
Durchsetzungsvermögen eines rezessiven (oberes Bild) und eines dominanten (unteres Bild) Allels unter der Voraussetzung, daß es einen Selektionsvorteil von 1 in 1000 (s = 0,001) hat. Auf der Ordinate sind die Allelhäufigkeiten zwischen 0 und 1eingetragen, auf der Abszisse die Zahl der Generationen von 1 bis 10000). (Nach T. O. SOLBRIG, 1970).
Wie groß ist die Wahrscheinlichkeit, daß sich eine Mutation in einer Population sich sexuell fortpflanzender Organismen durchsetzt? Unter Vernachlässigung der Selektion konnte R. A. FISHER zunächst zeigen, daß eine einmal aufgetretene Mutation, symbolisiert durch den Genotyp Aa in einer Population von Individuen des Genotyps AA, kaum eine Chance hat, sich durchzusetzen. Um das Allel a auf die Nachkommenschaft zu übertragen, muß sich das Aa-Individuum mit einem der AA-Individuen paaren. Die Wahrscheinlichkeit des Verlusts des Allels a gibt FISHUR mit e-1 pro Generation an. Die Verlustrate (Eliminierungsrate) wäre folglich durch eine Poissonverteilung zu beschreiben:
e-1 = 0,368.
Demnach wäre die Wahrscheinlichkeit für das Vorhandensein von a nach einer Generation
1 - 0,368 = 0,632
und nach einer weiteren Generation
e-(1- 0368) = 0,531
Verallgemeinernd heißt das, daß das Allel in 90 Prozent aller Fälle spätestens nach 15 Generationen verschwunden ist. Ein zufälliger Verlust oder Erwerb eines nichtadaptiven Allels in einer Population wird als (genetische) Drift bezeichnet. Anstatt durch Mutation kann ein solches Allel durch Genwanderung (Migration) von Individuen fremder Populationen in eine Population eingebracht worden sein. Der Genaustausch zwischen Populationen wird Genfluß genannt.
Mutationen trifft man in natürlichen Populationen sehr häufig an. Man schätzt, daß sie mit einer Häufigkeit von 10-5 bis 10-6 je Genort und Generation auftreten, wobei die Wahrscheinlichkeit des Auftretens von Genort zu Genort ganz beträchtlich schwanken kann. Bei einer durchschnittlichen Mutationsrate wäre die Austauschwahrscheinlichkeit eines Allels durch ein anderes, ihm gleichwertiges, außerordentlich gering. Eine Verringerung von z.B. p = 0,5 auf p = 0,1 (=Verringerung des Anteils von 50 auf 10 Prozent), würde 160 000 Generationen in Anspruch nehmen. Das zeigt, daß die Entstehung der Formenvielfalt, so wie sie sich uns in der Natur darstellt, nicht auf Akkumulation neutraler Mutationen beruhen kann. Die Situation ändert sich jedoch drastisch, wenn man einen Selektionsvorteil veranschlagt.
Ein vorteilhaftes dominantes Allel kann sich in einer Population sehr schnell ausbreiten, rezessive Allele haben trotz Selektionsvorteil ein nur geringes Durchsetzungsvermögen. Dabei spielt die Populationsgröße eine entscheidende Rolle, denn je kleiner eine Population ist, desto schneller kann sich eine günstige Mutation etablieren. Alle neuen Arten sind aus kleinen Ausgangspopulationen hervorgegangen, die E. MAYR (1942) Gründerpopulationen genannt hat. Inzucht fördert die Etablierung von Mutationen. Aber: durch Inzucht und in Gründerpopulationen werden nicht nur fördernde Mutationen angereichert, sondern ebenso auch nachteilige, und die Folge davon ist eine hohe Aussterbequote inzuchttreibender Populationen sowie von "Gründerpopulationen".

Durchsetzungsvermögen von Mutationen (gelb, rot, blau und grün) in großen und kleinen Populationen bei asexueller und sexueller Fortpflanzung. Die Kombination von Mutanten ist in den Bildern durch Mischfarben angezeigt. Einzelheiten s. nachfolgenden Text (Nach J. F. CROW und M. KIMURA, 1979).
Das vorgestellte Modell beruht auf der Annahme, daß Allele an drei Genorten (A, B, C) adaptiv sind und gleichzeitig auftreten. Nur bei sexueller Fortpflanzung kommen sie in kurzer Zeit, durch Rekombination vereint, gemeinsam zur Wirkung. Bei asexueller Fortpflanzung kann eine Addition von A und B erst dann erfolgen, wenn die Mutation zu B in einer Population A-tragender Individuen erneut auftritt (H. J. MULLER, 1932).
Jede natürliche Population besteht aus einer Vielzahl von Genotypen (reinen Linien im Sinne JOHANNSENs). Das heißt, daß die Selektion keineswegs auf alle Individuen gleichermaßen wirkt. Die Folge davon ist eine Veränderung der Populationsstruktur nach erfolgter Selektion. Rein formal lassen sich dabei mehrere Möglichkeiten voneinander unterscheiden, die, wie die folgende Abbildung zeigt, zu unterschiedlichen Ergebnissen führen.
Am einschneidendsten ist dabei die sogenannte disruptive Selektion, denn durch sie werden die Durchschnittsgenotypen (damit also die Mehrheit der Individuen einer Population) benachteiligt. Bewirkt wird dadurch eine Bevorzugung der extremen Genotypen, und die Bildung einer zweigipfligen Verteilung kann, wenn verschiedene andere Faktoren hinzukommen, zum Ersatz der ursprünglichen Population (Art) durch Entstehung von zwei neuen (Arten) führen.
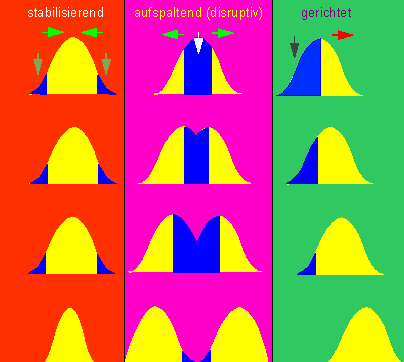
Drei Arten der Selektion. 1. (rotes Feld): stabilisierende Selektion. Die Umweltfaktoren favorisieren Individuen, die dem Populationsdurchschnitt entsprechen. In Verlauf der Zeit nimmt die Variabilität innerhalb der Population ab. 2. (violettes Feld): aufspaltende (disruptive) Selektion. Der Selektionsdruck richtet sich gegen den Populationsdurchschnitt. Die Extreme gewinnen an Vorteil. Die Population spaltet sich in zwei Teilpopulationen. 3. (grünes Feld): gerichtete Selektion. Der Selektionsdruck richtet sich nur gegen Individuen an der einen Seite der Verteilung. Die Kurve verschiebt sich daher. Dieser Typ ist in natürlichen Populationen der bei weitem häufigste (Nach O. T. SOLBRIG 1970, O. T. SOLBRIG, D. J. SOLBRIG, 1979).
|
|