
Schon kurz nach der Wiederentdeckung der MENDELschen Regeln beobachtete H. de VRIES neu auftretende Formen in Populationen von Oenothera. Sie zeichneten sich gegenüber den übrigen, weitverbreiteten Exemplaren (dem Wildtyp) durch andersartigen Wuchs, anders geformte Blätter sowie durch unterschiedliche Entwicklungsgeschwindigkeit aus. Die Änderungen des Phänotyps blieben auch in der Nachkommenschaft erhalten, stellten sich damit also als erblich heraus. Sie beruhten offensichtlich auf Veränderungen der Erbanlagen, ohne daß eine Ursache zu erkennen war. Derartige Erbänderungen nennt man Mutationen und Organismen, in denen sie zum Zuge kommen, Mutanten.
Unter dem Ausdruck "Wildtyp" versteht man ganz allgemein die in der Natur am häufigsten auftretenden Phänotypen (einer betreffenden Art). Unterscheidet sich eine Mutante in einem bestimmten Merkmal vom Wildtyp, so kennzeichnet man das mutierte Allel durch eine geeignete Abkürzung, das Wildtypallel durch +.
Den Mutationen stellt man die Modifikationen gegenüber. Der Begriff beschreibt umweltbedingte Änderungen des Phänotyps. Dazu ein Beispiel: E. S. ROBERTSON und I. C. ANDERSON (1961) charakterisierten eine Mutante vom Mais, die sich in rezessiv homozygotem Zustand (v/v) bei niedriger Temperatur (< 20 ºC) durch eine blasse Färbung der Blätter (virescent) auszeichnete.
Bei erhöhter Temperatur (37 ºC) glichen die Pflanzen dem Wildtyp. Die F1-Hybriden (+/v) waren dunkelgrün wie der Wildtyp), in der F2 wurde eine 3:1-Aufspaltung gefunden, als die Pflanzen bei 20 ºC kultiviert wurden. Erfolgte die Kultur ledoch bei 37 ºC, erschienen einheitlich aussehende, dunkelgrüne Pflanzen.
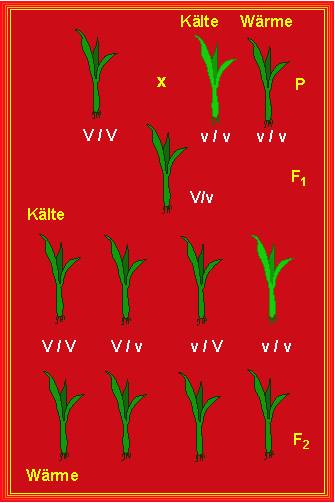
Dieser Fall veranschaulicht, daß die Ausbildung eines Phänotyps
durch Umweltfaktoren (hier die Temperatur) beeinflußt werden
kann.
Gerade bei Pflanzen sind Modifikationen an der Tagesordnung. Zu den bekannten Lehrbuchbeispielen gehört der Befund, daß die Blütenfarbe einer bestimmten Sorte von Primula sinensis bei Raumtemperatur rot, bei erhöhter Temperatur weiß erscheint. Mutanten, die erst bei erhöhter Temperatur erkennbar sind, nennt man temperatursensitive Mutanten.
Hochgebirgsformen und überhaupt Pflanzen, die einer intensiven
Strahlung ausgesetzt sind, zeigen gedrungenen Wuchs. Flachlandformen
und solche, die während ihres Wachstums geringen Lichtintensitäten
ausgesetzt waren, zeigen deutliches Streckungswachstum. Man weiß,
daß gerade die Entwicklung der Pflanzen in weit stärkerem
Umfang noch als die der Tiere durch externe Faktoren gesteuert
wird. Der Begriff Photomorphogenese
belegt, welch wichtige Rolle dabei dem Licht zukommt. Eine in
Dunkelheit aufwachsende Pflanze kann weder Blüten noch Chloroplasten
bilden, obwohl die dazu nötigen Erbanlagen vorhanden sind.
Um jene zu aktivieren, bedarf es des entsprechenden externen Signals.
Damit wird deutlich, daß die Entwicklung einer Pflanze (und
die anderer Organismen) durch ein Wechselspiel zwischen Erbanlagen
und Umweltfaktoren gesteuert wird.
Es gehört heute zu den akzeptierten Fakten, daß die
Umwelt den Phänotyp beeinflußt, auf den Genotyp jedoch
keinen direkten, präziser ausgedrückt, richtenden Einfluß
ausübt.
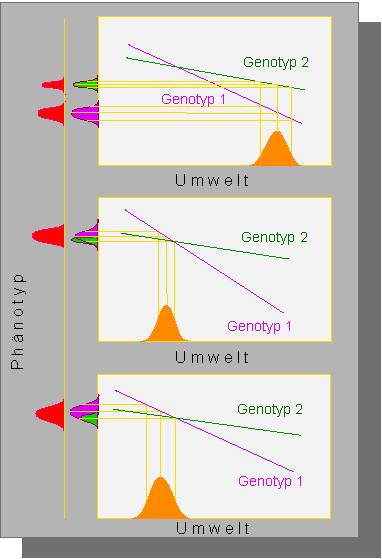
Die Auswirkungen von Umwelteinflüssen auf die Modifikation von Phänotypen zweier unterschiedlicher Genotypen.
Es gibt jedoch eine Anzahl von Chemikalien und physikalischen Faktoren, die Mutationen auslösen und damit die Mutationshäufigkeit erhöhen.
Zu den physikalischen Faktoren gehört in erster Linie die ionisierende Strahlung. 1927 wies J. H. MULLER nach, daß Röntgenstrahlung die Mutationsrate bei Drosophila heraufsetzt. Nahezu gleichzeitig wurden Mutanten auch bei verschiedenen Pflanzen induziert: Nicotiana tabacum (T. H. GOODSPEED), Datura stramonium (C. S. GAGER und A. F. BLAKESLEE, 1927) Zea mays und Hordeum vulgare (L. J. STADLER).
Die Mutagene wirken auf den Organismus fast ausnahmslos schädigend, ihre Wirkung ist zeit- und dosisabhängig, oft letal. Unter den wenigen Überlebenden ist die Mutationsrate gegenüber der entsprechenden Kontrolle drastisch erhöht.
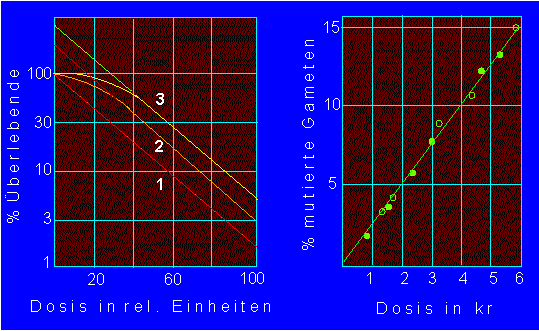
Linkes Diagramm: Inaktivierungskinetiken. Die drei Funktionen entsprechen (von unten nach oben) einer Eintreffer-, Zweitreffer- und Dreitrefferkinetik. Die Anzahl der notwendigen Treffer läßt sich aus dem Diagramm ablesen, wenn man den linearen Bereich der Kurven nach "rückwärts" extrapoliert (gestrichelt dargestellt). Der Schnittpunkt mit der Ordinate gibt die Zahl der Treffer an, z.B.300:100= 3,
Rechtes Diagramm: Mutationsauslösung bei Drosophila durch ionisierende Strahlung. Die unterschiedlichen Symbole im Diagramm beziehen sich auf unterschiedliche Strahlenquellen. (Nach K. G. ZIMMER und TIMOFEEFF-RESSOVSKY, 1942).
Doch nur in den allerseltensten Fällen entsteht durch ein mutagenes Agens (= Mutagen) eine Mutante, die gegenüber dem Wildtyp vorteilhaftere Eigenschaften aufweist oder eine höhere Lebenserwartung hat.
Der Versuch, durch Bestrahlung ertragreichere oder resistentere Sorten zu züchten, erwies sich trotz eines großen finanziellen Aufwands weitgehend als Fehlschlag. Anders sieht die Situation bei Mikroorganismen aus. Wegen ihrer hohen Vermehrungsrate ist es relativ einfach, auch noch so selten auftretende vorteilhafte Formen zu isolieren.
Nahezu alle in der industriellen Mikrobiologie verwendeten Stämme zur Produktion verschiedenartigster Stoffe (von der Zitronensäure bis zu Proteinen und Antibiotika) sind durch Mutagene erzeugt worden. Diese Stämme finden unter kontrollierten Laborbedingungen ideale Lebensbedingungen, haben aber kaum eine Chance, sich unter natürlichen Bedingungen gegen den jeweili'en Wildtyp durchzusetzen.
In der Evolutionsforschung spricht man von Fitneß und beschreibt damit den relativen Fortpflanzungserfolg eines Individuums. Die Fitneß von Mutanten ist meist sehr gering.
Mutagene können auf unterschiedliche Weise wirken, Mutationen daher verschiedenen Typen angehören. Rein formal unterscheidet man zwischen den:
![]() Punktmutationen (= Genmutationen). Dabei sind einzelne Gene
betroffen.
Punktmutationen (= Genmutationen). Dabei sind einzelne Gene
betroffen.
![]() Chromosomenmutationen. Dabei werden Änderungen im Chromosomensatz
sichtbar, und
Chromosomenmutationen. Dabei werden Änderungen im Chromosomensatz
sichtbar, und
![]() Genommutationen. Dabei werden Chromosomensätze en
bloc verändert, z.B. verdoppelt.
Genommutationen. Dabei werden Chromosomensätze en
bloc verändert, z.B. verdoppelt.
Obwohl sich die Produktion pflanzlicher Mutanten wirtschaftlich nicht rentiert hat, erwies sie sich als das wohl wirkungsvollste experimentelle Instrument moderner Grundlagenforschung. Die Analyse der verschiedensten Mutanten gab Aufschlüsse über die Struktur und Funktion von Erbanlagen sowie über ihren Einfluß auf die Entwicklung eines Individuums.
E. BAUR definierte zusammenfassend in seinem Lehrbuch der Vererbungslehre Modifikationen, Mutationen und Kombinationen (Bastardbildung) wie folgt:
Modifikationen: nicht erbliche Verschiedenheiten zwischen den Individuen einer Population (oder Art), verursacht durch diverse Außeneinwirkungen, wie Licht, Wärme, Ernährung usw., die die einzelnen Individuen ungleich beeinflussen.
Mutationen: erbliche Verschiedenheiten zwischen den Eltern und ihren Nachkommen, auch den vegetativ entstandenen, welche nicht auf Bastardbildung beruhen, sondern andere Ursachen haben.
Kombinationen: erbliche Verschiedenheiten zwischen den Individuen einer Population und auch zwischen den Nachkommen eines Elternpaares, verursacht durch Bastardspaltung und Neukombination der Erbanlagen.
Diese drei Kategorien sind dem bloßen Aussehen nach nicht zu unterscheiden. Worauf das "Variieren" irgendeines Individuums beruht, ist meistens nur durch sorgfältige Vererbungsversuche feststellbar. Nimmt man solche Versuche als Kriterium für die Unterscheidung, dann ist die Trennung der drei Kategorien fast immer sicher durchführbar.
|
|