
Wie eben dargelegt, durchlief die Vererbungsforschung im ausgehenden 18. und im beginnenden 19. Jahrhundert eine sehr arbeitsreiche Phase, in der eine Vielzahl an Beobachtungen gesammelt wurde und zahlreiche Schlüsse gezogen wurden. Es fehlte aber der entscheidende Durchbruch. In der Systematik gelang ein solcher Carl v. LINNÉ , in der Genetik Gregor MENDEL.
Johann MENDEL (der Vorname Gregor wurde ihm nach seinem Eintritt ins Kloster verliehen), geb. 1822 in Heinzendorf (im deutschen Teil des damals österreichischen Schlesiens), Abt in Brünn, gest. 1884, publizierte 1866 eine zwar kleine aber inhaltsschwere Schrift mit dem Titel
die zum grundlegenden Werk und Ausgangspunkt der modernen Genetik wurde.
:
Über seine Versuchspflanzen schrieb er:
"Die Auswahl der Pflanzengruppe muß mit möglichster Vorsicht geschehen, wenn man nicht im vorhinein allen Erfolg in Frage stellen will.
Die Versuchspflanzen müssen notwendig
Konstant differierende Merkmale besitzen.
Die Hybriden derselben müssen während der Blütezeit vor der Einwirkung jedes fremdartigen Pollens geschützt sein oder leicht geschützt werden können.
Dürfen die Hybriden und ihre Nachkommen in den aufeinanderfolgenden Generationen keine merkliche Störung der Fruchtbarkeit erlangen. Fälschungen durch fremden Pollen, wenn solche im Verlauf des Versuchs vorkämen, müßten zu ganz irrigen Ansichten führen. Verminderte Fruchtbarkeit oder gänzliche Sterilität einzelner Formen, wie sie unter den Nachkommen vieler Hybriden vorkommen, würden die Versuche erschweren oder ganz vereiteln."
Er wählte Pisum sativum als Versuchspflanze und konzentrierte sich auf die Analyse von sieben Merkmalspaaren. Die Wahl des Objekts war auch insofern glücklich, als die Kulturform in der Regel ein Selbstbestäuber ist, die einzelnen Linien (Sorten) daher einen hohen Inzuchtgrad aufweisen und er deshalb auf reine Linien zurückgreifen konnte. MENDEL gelangte zu den folgenden Ergebnissen und Aussagen:
Dazu noch einige Bemerkungen zur modernen Terminologie. Die Elterngeneration wird mit P (= Parentalgeneration) bezeichnet. Durch Bastardierung erhält man die erste Filialgeneration (Tochtergeneration): F1. Kreuzt man die Bastarde untereinander, kommt man zur F2, eine Generation weiter zur F3 usw. Der Begriff Uniformitätsregel beschreibt die Homogenität der Individuen in der F1. Dabei kann (wie bei allen von MENDEL an der Erbse untersuchten Merkmalspaaren) eines der Merkmale dominant über das andere sein (dominant-rezessiver Erbgang). Alle Individuen der F1 sind dann durch das dominante Merkmal ausgezeichnet. Es gibt aber auch Fälle, bei denen die Merkmalsausprägung in der F1 eine Zwischenstellung zwischen den Merkmalen der beiden Eltern einnimmt. In diesen Fällen spricht man von intermediärem Erbgang.
Reziprozitätsregel bedeutet, daß es bei der Kreuzung zweier Pflanzen (mit Unterschieden in einem Merkmalspaar) belanglos ist, ob das Merkmal vom Vater oder von der Mutter (oder umgekehrt = reziprok) stammt.:
Hierzu ein Beispiel: (weiblich) Samen rund x (männlich) Samen kantig ergibt F1 Individuen (weiblich und männlich), deren Samen rund sind, da rund über kantig dominant ist. Zum gleichen Ergebnis kommt man durch Verwendung des folgenden Elternpaares: (weiblich) Samen kantig x (männlich) Samen rund. Es hat sich durchgesetzt, in Kreuzungsexperimenten den weiblichen Elter (Elter = Singular von Eltern) an erster, den männlichen an zweiter Stelle zu schreiben.
| Verhältniszahlen für differierende
Merkmale in der "ersten Generation der Hybriden": F2 |
|||
|---|---|---|---|
| Merkmalspaar | Individuen |
||
| 1. | Samen: rund-kantig | ||
| 2. | Kotyledonen: gelb-grün | ||
| 3. | Samenschale: grau-weiß | ||
| 4. | Hülse: einfach gewölbt-eingeschnürt | ||
| 5. | Unreife Hülse: grün-gelb | ||
| 6. | Blüte: achsenständig-endständig | ||
| 7. | Blütenachse: lang-kurz | ||
Wurden nunmehr die Hybriden (Bastarde) untereinander gekreuzt, traten in der darauffolgenden Generation (= F2) die rezessiven Formen wieder auf. Aus den experimentell ermittelten Verhältniszahlen extrapolierte Mendel auf das Verhältnis 3:1.
Daraus folgerte er:
"Da die Glieder der ersten Hybridgeneration (Anm: F2) unmittelbar aus den Samen der Hybriden hervorgehen, wird es nun ersichtlich, daß die Hybriden je zweier differierender Merkmale Samen bilden, von denen die eine Hälfte wieder die Hybridform entwickelt, während die andere Pflanzen gibt, welche konstant bleiben und zu gleichen Teilen den dominierenden und den rezessiven Charakter erhalten."
MENDEL ging davon aus, daß sowohl die Pollenzellen als auch die Keimzellen (=Eizellen) Anlagen tragen. Damit hatte er eine weitere wichtige Erkenntnis gewonnen; es werden nämlich nicht die Eigenschaften an sich, sondern die Anlagen dafür vererbt.
Da es vom Zufall abhängt, welche Pollen- und Keimzellen (A, a) sich miteinander verbinden, ergeben sich in der Nachkommenschaft die Kombinationen:
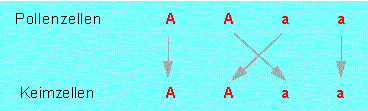
Diese Gesetzmäßigkeit bringen wir heute durch die 2. MENDELsche Regel (=Spaltungsregel) zum Ausdruck: Die F2-Individuen sind untereinander nicht alle gleich, vielmehr werden unterschiedliche Erscheinungsformen sichtbar. Stets treten die Merkmale der Ausgangsformen (der Parentalgeneration: P) in bestimmten Zahlenverhältnissen wieder auf. Je nachdem, ob ein dominant-rezessiver oder ein intermediärer Erbgang vorliegt, erhält man eine Aufspaltung von 3:1 oder 1:2:1.

Intermediärer Erbgang der Blütenfarben von Mirabilis jalapa. AA-Genotypen haben rote, Aa-Genotypen rosa und aa-Genotypen weiße Blüten (Nach C. CORRENS, 1902)
Beim dominant-rezessiven Erbgang sieht man den Individuen, die sich durch das dominante Merkmal auszeichnen, nicht an, ob es in nachfolgenden Generationen erhalten bleibt oder aufspaltet.
Zur Klärung des Sachverhalts einige formale Betrachtungen: Eine Merkmalsanlage, z.B. Samenform, Kotyledonenfarbe, Samenschalenfarbe wird (einem Vorschlag von BATESON [1909] folgend) Gen genannt. Die Zustandsform, in der ein Gen vorliegt, z.B. bei der Samenform rund oder kantig, bezeichnet man als Allel. Jedes Individuum erhält pro Gen ein Allel von der Mutter, ein weiteres vom Vater. Beide Allele können gleichartig, sie können aber auch verschieden sein. Im ersten Fall wäre das Individuum homozygot (die Aussage bezieht sich immer nur auf das zu untersuchende Merkmal), im zweiten heterozygot. Bereits MENDEL kennzeichnete Gene (er nannte sie seinerzeit Anlagen) durch Buchstaben (s.o.). Auch hier gibt es heute nach internationalen Konventionen Regeln, die die Schreibweise solcher Abkürzungen festlegen. Es ist aber leider so, daß sich in den einzelnen Teildisziplinen der Genetik unterschiedliche Terminologien eingebürgert haben. Drosophilagenetiker verwenden andere Bezeichnungen als etwa die Bakteriengenetiker.
Zunächst aber zu einer Grundregel: Ein dominantes Allel wird durch einen Großbuchstaben, ein rezessives durch den entsprechenden kleinen Buchstaben gekennzeichnet. Kreuzt man beispielsweise eine rotblühende Pflanze (rot sei dominant) mit einer weißblühenden (weiß demnach rezessiv), darf man nicht mit den Abkürzungen r (= rot) und w (= weiß) operieren, sondern muß die rotblühende Form mit R, die weißblühende mit r kennzeichnen. Da in einem Individuum pro Gen zwei Allele vorliegen, gibt es die beiden homozygoten Formen RR und rr, zum anderen die heterozygote(n) Rr (= rR).
Bei dominant-rezessivem Erbgang lassen sich die Homozygoten (RR) unter den Individuen mit dominanter Merkmalsausprägung nicht von den Heterozygoten (Rr) unterscheiden. Um eine Entscheidung herbeizuführen, werden einzelne Individuen mit dem rezessiven Elter (rr) gekreuzt. Dabei hat man es mit einer Rückkreuzung zu tun. Man erhält, von Homozygoten (RR x rr) ausgehend, in der darauffolgenden Generation ausschließlich Rr (alle Individuen sind uniform), während man von Heterozygoten (Rr x rr) ausgehend, zu je 50 Prozent Rr und rr kommt.
Da man einem Individuum unter den hier beschriebenen Voraussetzungen zunächst nicht ansehen kann, ob das entsprechende Gen in homozygotem oder heterozygotem Zustand (RR oder Rr) vorliegt, unterscheidet man zwischen Phänotyp (= Erscheinungsform) und dem Genotyp (= der allelen Zusammensetzung). Einem bestimmten Phänotyp können demnach mehrere (hier zwei) Genotypen zugrunde liegen.
Kombination mehrerer Merkmalspaare. - MENDELs Beispiel:
In der Fl hatten alle Individuen runde Samen und gelbe Kotyledonen, in der F2 spalteten sie wie folgt auf:
315 runde Samen und gelbe Kotyledonen
101 kantige Samen und gelbe Kotyledonen
38 kantige Samen und grüne Kotyledonen
108 runde Samen und grüne Kotyledonen
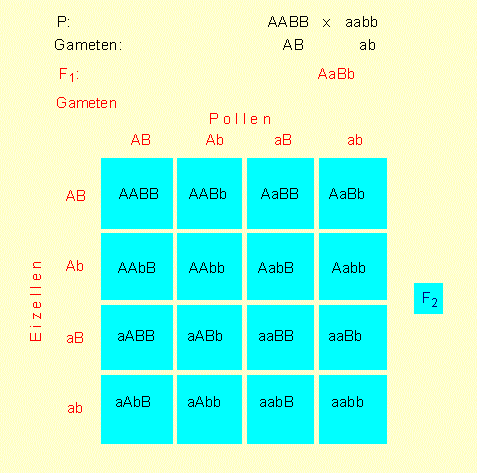
PUNNETT-Square: Schema zur Wiedergabe der Genotypen in der P-, F1- und F2-Generation. Beispiel für einen dihybriden Erbgang. Diese Art der Darstellung wurde zu Beginn des Jahrhunderts durch den britischen Genetiker R. C. PUNNETT eingeführt.
Daraus ergibt sich, daß die Merkmale unabhängig voneinander vererbt werden. In der F2 treten die Genotypen im Zahlenverhältnis 9:3:3:1 auf. Damit wären wir bei der 3. MENDELschen Regel (Unabhängigkeitsregel oder Regel von der Neukombination verschiedener Genpaare). Die Unabhängigkeitsregel beinhaltet zwangsläufig, daß Genkombinationen neu entstehen können, die ursprünglich nicht vorhanden waren, nämlich:
runde Samen und grüne Kotyledonen und
kantige Samen und gelbe Kotyledonen.
Untersucht man nur ein Merkmal (wie bei der Besprechung der 1. und 2. MENDELschen Regel angenommen), spricht man von monohybridem Erbgang. Kommen weitere Merkmale (Gene) hinzu, von dihybridem, trihybridem... polyhybridem Erbgang.
Ohne mathematische Ableitung sind Mendels Schlußfolgerungen nur schwer zu verstehen. Übungsaufgaben sind daher unumgänglich: Mendels Experiment kann man hier selbst durchführen:
|
|